

Epac/Rap and PKA are novel mechanisms of ANP-induced Rac-mediated pulmonary endothelial barrier protection
- PMID: 18064650
- PMCID: PMC3652875
- DOI: 10.1002/jcp.21354
Epac/Rap and PKA are novel mechanisms of ANP-induced Rac-mediated pulmonary endothelial barrier protection
Abstract
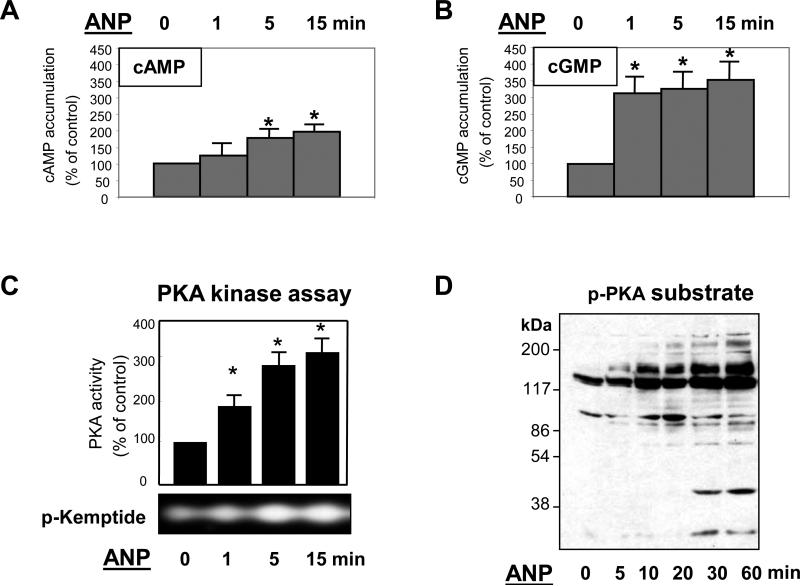
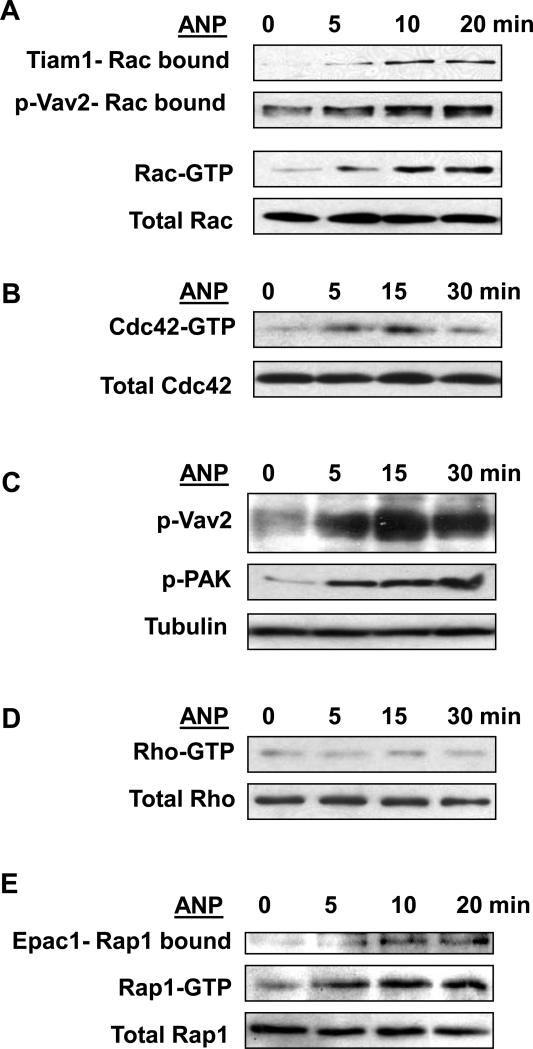
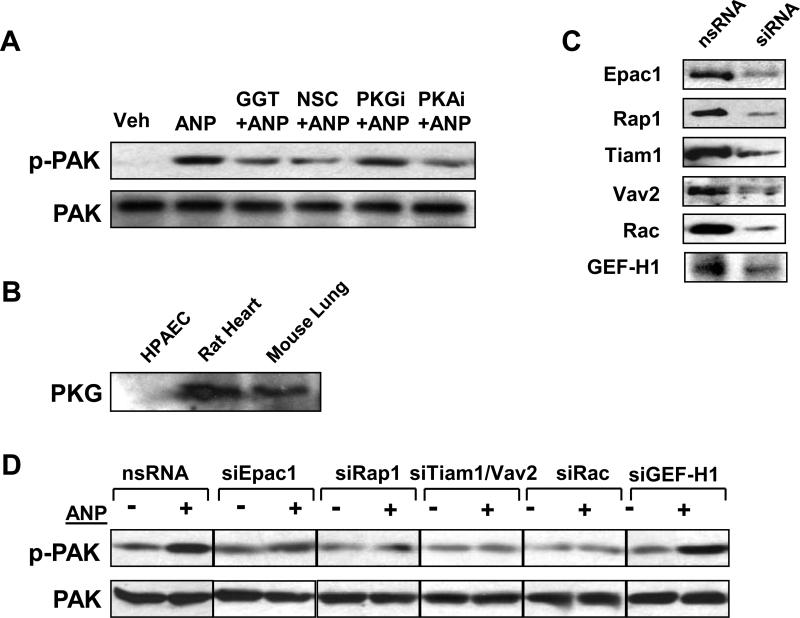
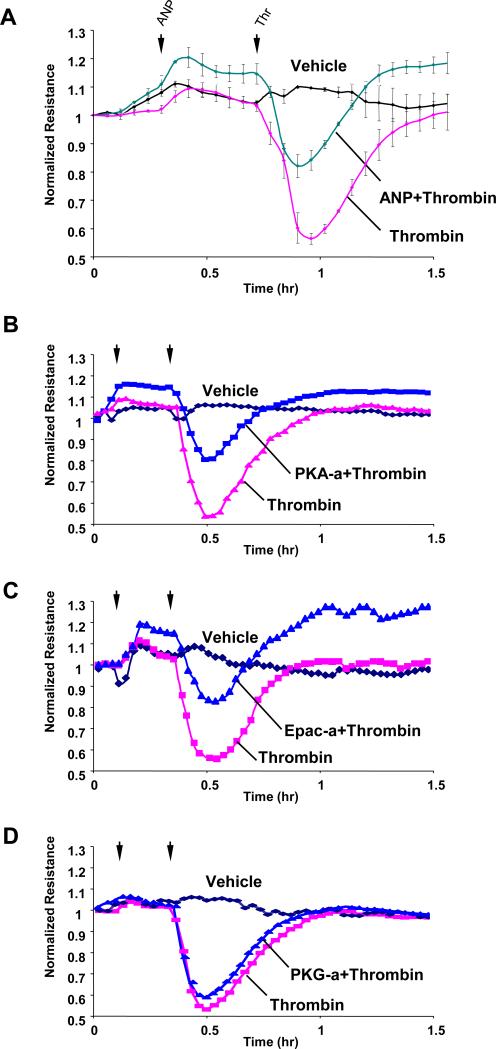
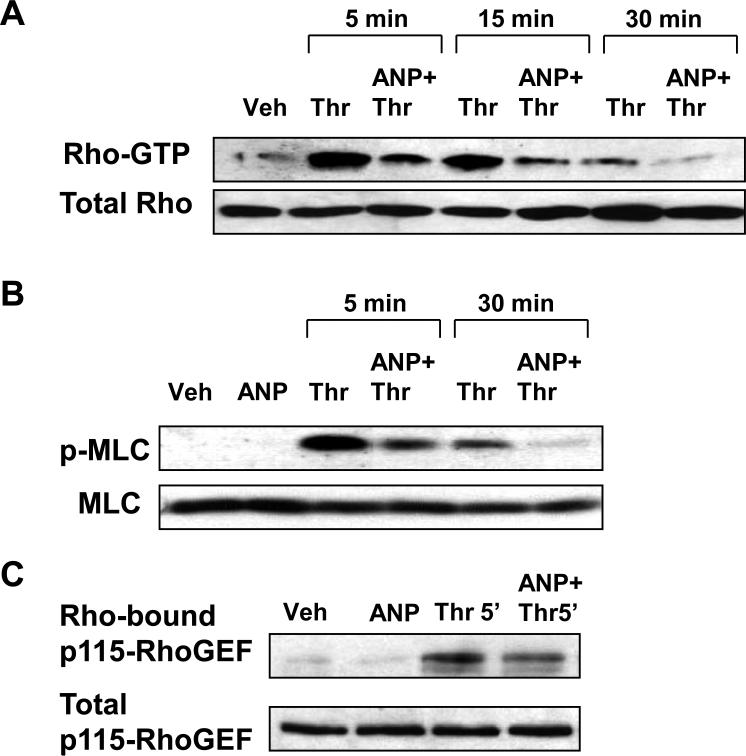
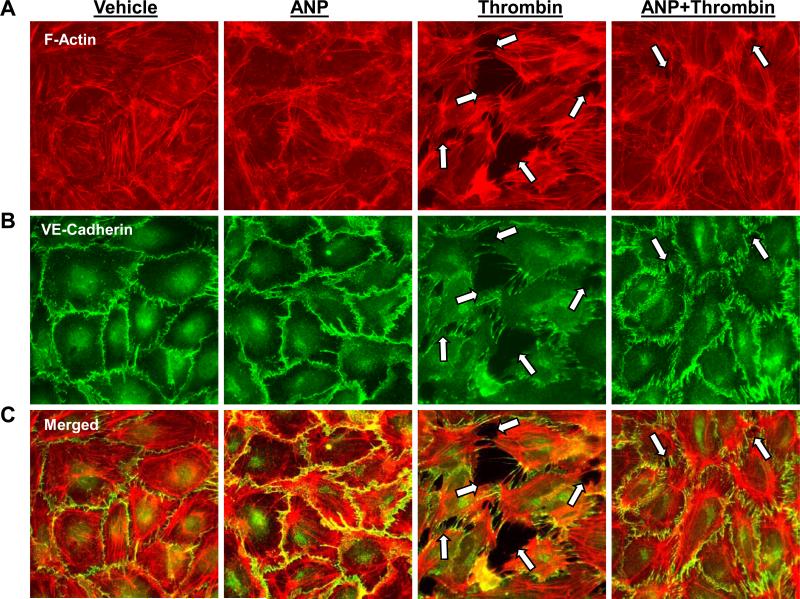
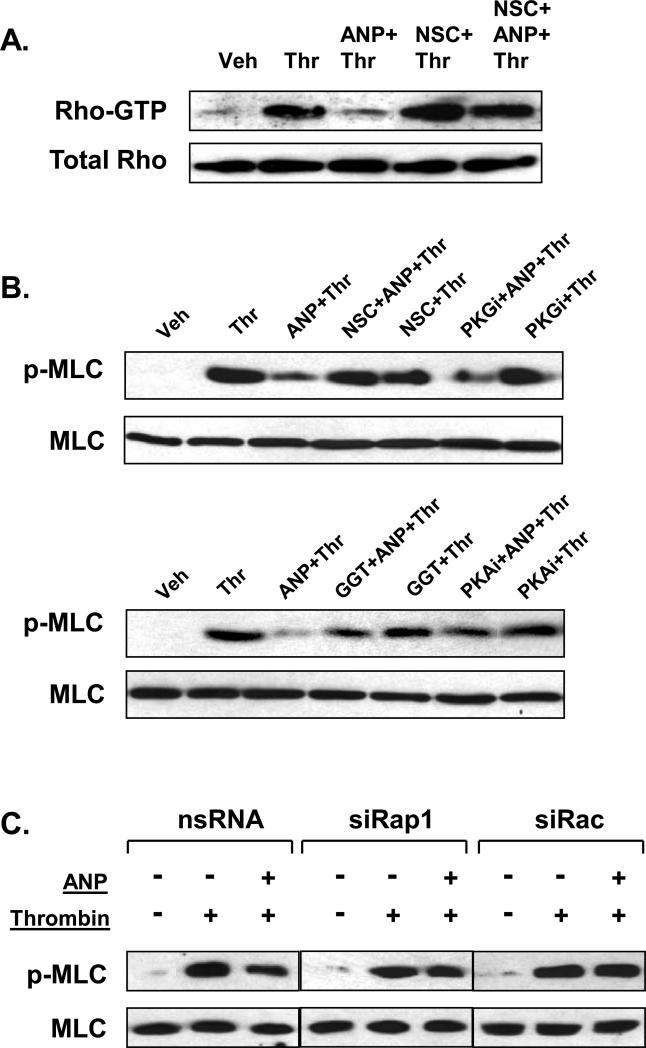
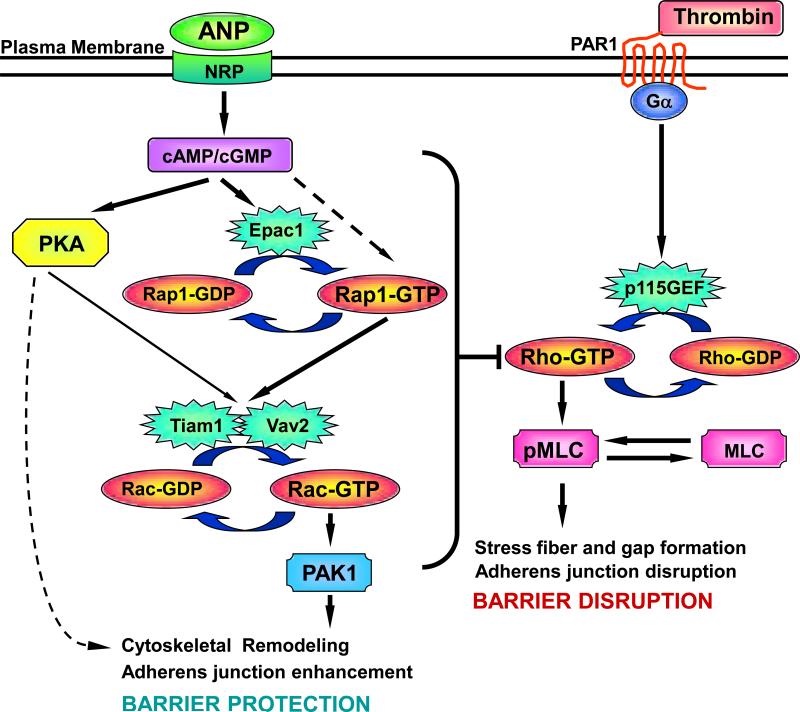
Acute lung injury, sepsis, lung inflammation, and ventilator-induced lung injury are life-threatening conditions associated with lung vascular barrier dysfunction, which may lead to pulmonary edema. Increased levels of atrial natriuretic peptide (ANP) in lung circulation reported in these pathologies suggest its potential role in the modulation of lung injury. Besides well recognized physiological effects on vascular tone, plasma volume, and renal function, ANP may exhibit protective effects in models of lung vascular endothelial cell (EC) barrier dysfunction. However, the molecular mechanisms of ANP protective effects are not well understood. The recently described cAMP-dependent guanine nucleotide exchange factor (GEF) Epac activates small GTPase Rap1, which results in activation of small GTPase Rac-specific GEFs Tiam1 and Vav2 and Rac-mediated EC barrier protective responses. Our results show that ANP stimulated protein kinase A and the Epac/Rap1/Tiam/Vav/Rac cascade dramatically attenuated thrombin-induced pulmonary EC permeability and the disruption of EC monolayer integrity. Using pharmacological and molecular activation and inhibition of cAMP-and cGMP-dependent protein kinases (PKA and PKG), Epac, Rap1, Tiam1, Vav2, and Rac we linked ANP-mediated protective effects to the activation of Epac/Rap and PKA signaling cascades, which dramatically inhibited the Rho pathway of thrombin-induced EC hyper-permeability. These results suggest a novel mechanism of ANP protective effects against agonist-induced pulmonary EC barrier dysfunction via inhibition of Rho signaling by Epac/Rap1-Rac and PKA signaling cascades.
(c) 2007 Wiley-Liss, Inc.
Figures
References
-
- Aghazadeh B, Lowry WE, Huang XY, Rosen MK. Structural basis for relief of autoinhibition of the Dbl homology domain of proto-oncogene Vav by tyrosine phosphorylation. Cell. 2000;102(5):625–633. - PubMed
-
- Ahluwalia A, MacAllister RJ, Hobbs AJ. Vascular actions of natriuretic peptides. Cyclic GMP-dependent and -independent mechanisms. Basic Res Cardiol. 2004;99(2):83–89. - PubMed
-
- Baxter GF. The natriuretic peptides. Basic Res Cardiol. 2004;99(2):71–75. - PubMed
-
- Birukov KG, Bochkov VN, Birukova AA, Kawkitinarong K, Rios A, Leitner A, Verin AD, Bokoch GM, Leitinger N, Garcia JG. Epoxycyclopentenone-containing oxidized phospholipids restore endothelial barrier function via Cdc42 and Rac. Circ Res. 2004;95(9):892–901. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
