

Structure of [NiFe] hydrogenase maturation protein HypE from Escherichia coli and its interaction with HypF
- PMID: 18065529
- PMCID: PMC2238214
- DOI: 10.1128/JB.01610-07
Structure of [NiFe] hydrogenase maturation protein HypE from Escherichia coli and its interaction with HypF
Abstract
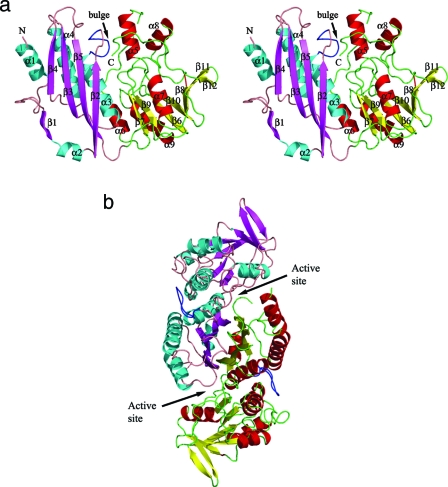
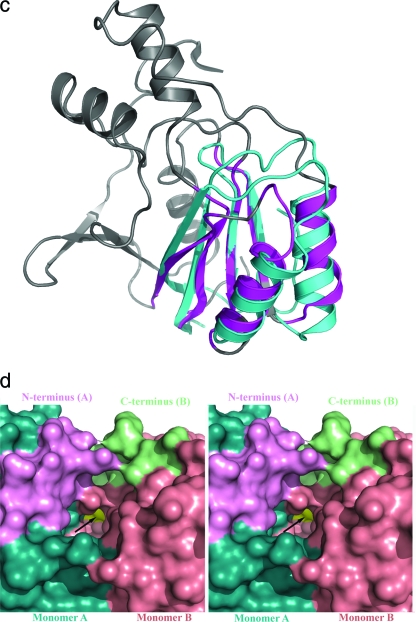
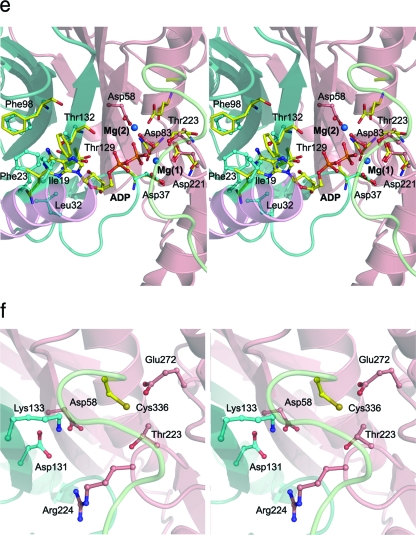
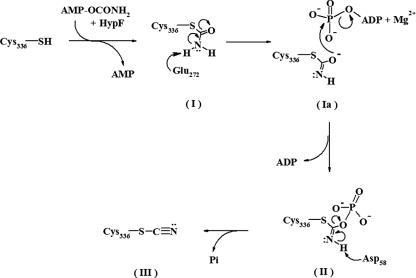
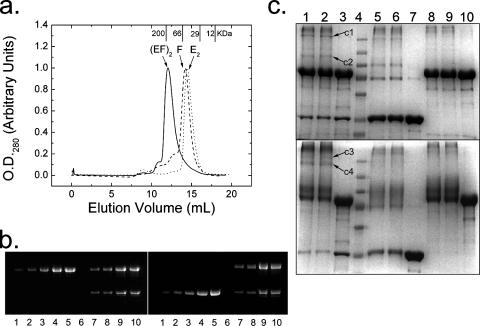
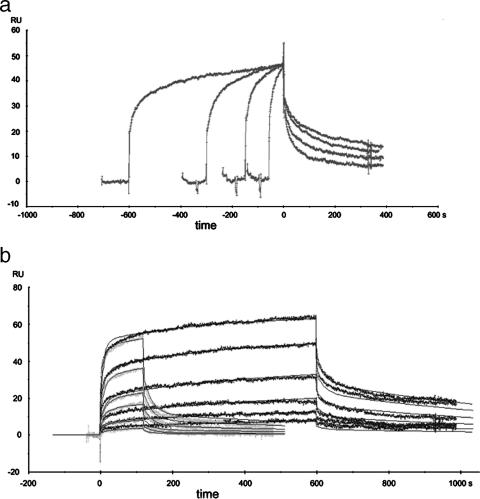
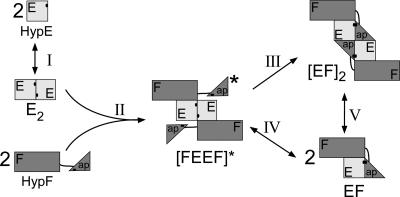
Hydrogenases are enzymes involved in hydrogen metabolism, utilizing H2 as an electron source. [NiFe] hydrogenases are heterodimeric Fe-S proteins, with a large subunit containing the reaction center involving Fe and Ni metal ions and a small subunit containing one or more Fe-S clusters. Maturation of the [NiFe] hydrogenase involves assembly of nonproteinaceous ligands on the large subunit by accessory proteins encoded by the hyp operon. HypE is an essential accessory protein and participates in the synthesis of two cyano groups found in the large subunit. We report the crystal structure of Escherichia coli HypE at 2.0-A resolution. HypE exhibits a fold similar to that of PurM and ThiL and forms dimers. The C-terminal catalytically essential Cys336 is internalized at the dimer interface between the N- and C-terminal domains. A mechanism for dehydration of the thiocarbamate to the thiocyanate is proposed, involving Asp83 and Glu272. The interactions of HypE and HypF were characterized in detail by surface plasmon resonance and isothermal titration calorimetry, revealing a Kd (dissociation constant) of approximately 400 nM. The stoichiometry and molecular weights of the complex were verified by size exclusion chromatography and gel scanning densitometry. These experiments reveal that HypE and HypF associate to form a stoichiometric, hetero-oligomeric complex predominantly consisting of a [EF]2 heterotetramer which exists in a dynamic equilibrium with the EF heterodimer. The surface plasmon resonance results indicate that a conformational change occurs upon heterodimerization which facilitates formation of a productive complex as part of the carbamate transfer reaction.
Figures
Similar articles
-
Structural basis for the reaction mechanism of S-carbamoylation of HypE by HypF in the maturation of [NiFe]-hydrogenases.J Biol Chem. 2012 Aug 17;287(34):28409-19. doi: 10.1074/jbc.M112.387134. Epub 2012 Jun 27. J Biol Chem. 2012. PMID: 22740694 Free PMC article.
-
Structure of hydrogenase maturation protein HypF with reaction intermediates shows two active sites.Structure. 2011 Dec 7;19(12):1773-83. doi: 10.1016/j.str.2011.09.023. Structure. 2011. PMID: 22153500
-
Proteolytic cleavage orchestrates cofactor insertion and protein assembly in [NiFe]-hydrogenase biosynthesis.J Biol Chem. 2017 Jul 14;292(28):11670-11681. doi: 10.1074/jbc.M117.788125. Epub 2017 May 24. J Biol Chem. 2017. PMID: 28539366 Free PMC article.
-
Structural Insight into [NiFe] Hydrogenase Maturation by Transient Complexes between Hyp Proteins.Acc Chem Res. 2020 Apr 21;53(4):875-886. doi: 10.1021/acs.accounts.0c00022. Epub 2020 Mar 31. Acc Chem Res. 2020. PMID: 32227866 Review.
-
Maturation of [NiFe]-hydrogenases in Escherichia coli.Biometals. 2007 Jun;20(3-4):565-78. doi: 10.1007/s10534-006-9048-5. Epub 2007 Jan 11. Biometals. 2007. PMID: 17216401 Review.
Cited by
-
Crystal structures of the carbamoylated and cyanated forms of HypE for [NiFe] hydrogenase maturation.Proc Natl Acad Sci U S A. 2013 Dec 17;110(51):20485-90. doi: 10.1073/pnas.1313620110. Epub 2013 Dec 2. Proc Natl Acad Sci U S A. 2013. PMID: 24297906 Free PMC article.
-
Anaerobic Formate and Hydrogen Metabolism.EcoSal Plus. 2016 Oct;7(1):10.1128/ecosalplus.ESP-0011-2016. doi: 10.1128/ecosalplus.ESP-0011-2016. EcoSal Plus. 2016. PMID: 27735784 Free PMC article. Review.
-
Metabolic deficiences revealed in the biotechnologically important model bacterium Escherichia coli BL21(DE3).PLoS One. 2011;6(8):e22830. doi: 10.1371/journal.pone.0022830. Epub 2011 Aug 3. PLoS One. 2011. PMID: 21826210 Free PMC article.
-
Structural basis for the reaction mechanism of S-carbamoylation of HypE by HypF in the maturation of [NiFe]-hydrogenases.J Biol Chem. 2012 Aug 17;287(34):28409-19. doi: 10.1074/jbc.M112.387134. Epub 2012 Jun 27. J Biol Chem. 2012. PMID: 22740694 Free PMC article.
-
Identification of an Isothiocyanate on the HypEF Complex Suggests a Route for Efficient Cyanyl-Group Channeling during [NiFe]-Hydrogenase Cofactor Generation.PLoS One. 2015 Jul 17;10(7):e0133118. doi: 10.1371/journal.pone.0133118. eCollection 2015. PLoS One. 2015. PMID: 26186649 Free PMC article.
References
-
- Anand, R., A. A. Hoskins, J. Stubbe, and S. E. Ealick. 2004. Domain organization of Salmonella typhimurium formylglycinamide ribonucleotide amidotransferase revealed by X-ray crystallography. Biochemistry 4310328-10342. - PubMed
-
- Andrews, S. C., B. C. Berks, J. McClay, A. Ambler, M. A. Quail, P. Golby, and J. R. Guest. 1997. A 12-cistron Escherichia coli operon (hyf) encoding a putative proton-translocating formate hydrogenlyase system. Microbiology 1433633-3647. - PubMed
-
- Blokesch, M., S. P. Albracht, B. F. Matzanke, N. M. Drapal, A. Jacobi, and A. Bock. 2004. The complex between hydrogenase-maturation proteins HypC and HypD is an intermediate in the supply of cyanide to the active site iron of [NiFe]-hydrogenases. J. Mol. Biol. 344155-167. - PubMed
-
- Blokesch, M., A. Paschos, A. Bauer, S. Reissmann, N. Drapal, and A. Bock. 2004. Analysis of the transcarbamoylation-dehydration reaction catalyzed by the hydrogenase maturation proteins HypF and HypE. Eur. J. Biochem. 2713428-3436. - PubMed
-
- Bock, A., P. W. King, M. Blokesch, and M. C. Posewitz. 2006. Maturation of hydrogenases. Adv. Microb. Physiol. 511-71. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
