

Genomic rearrangements and transcriptional analysis of the spliced leader-associated retrotransposon in RNA interference-deficient Trypanosoma brucei
- PMID: 18067542
- PMCID: PMC2610267
- DOI: 10.1111/j.1365-2958.2007.06057.x
Genomic rearrangements and transcriptional analysis of the spliced leader-associated retrotransposon in RNA interference-deficient Trypanosoma brucei
Abstract
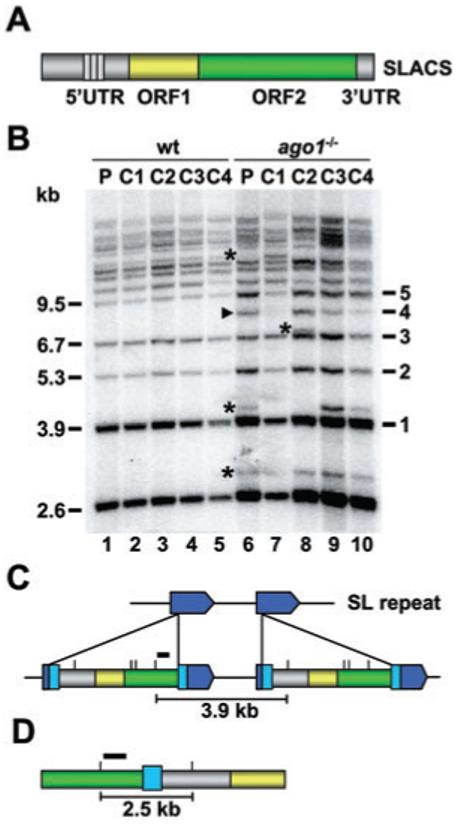
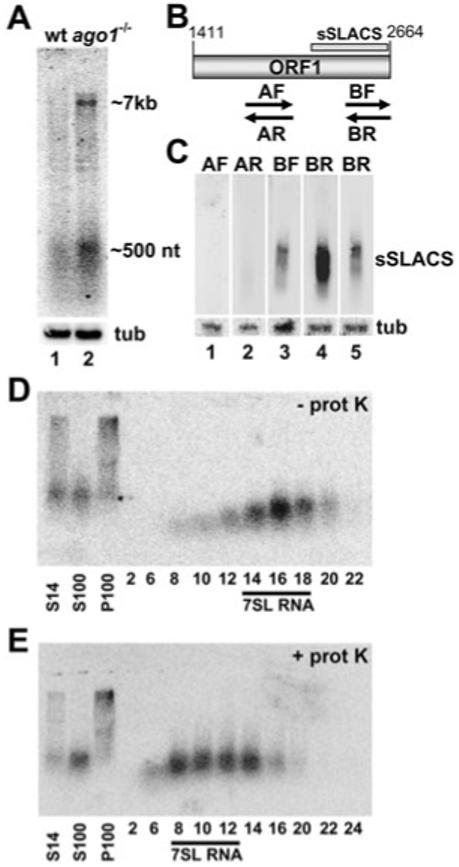
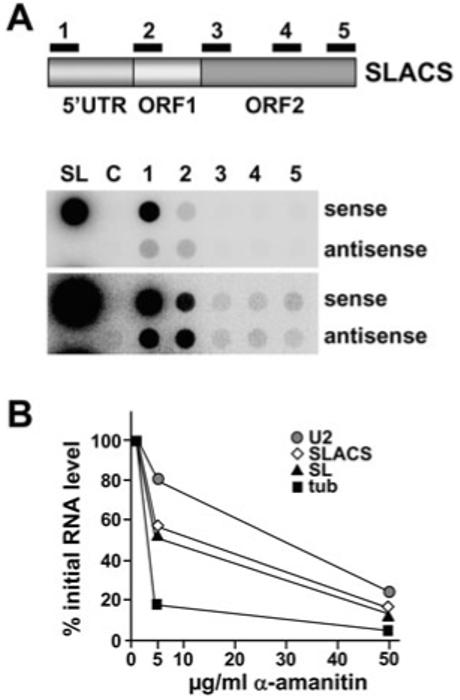
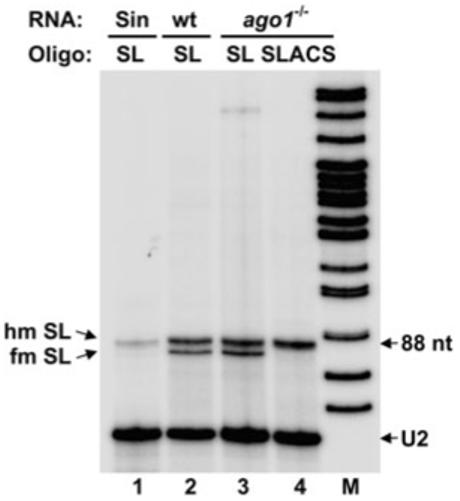
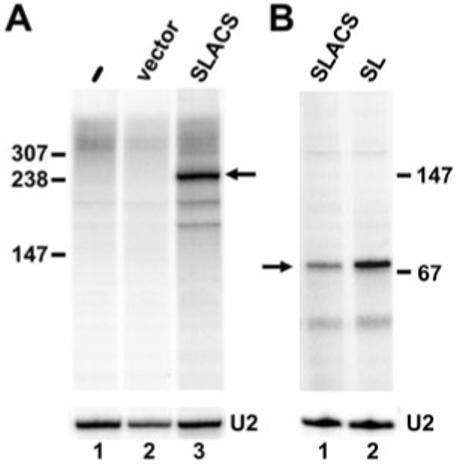
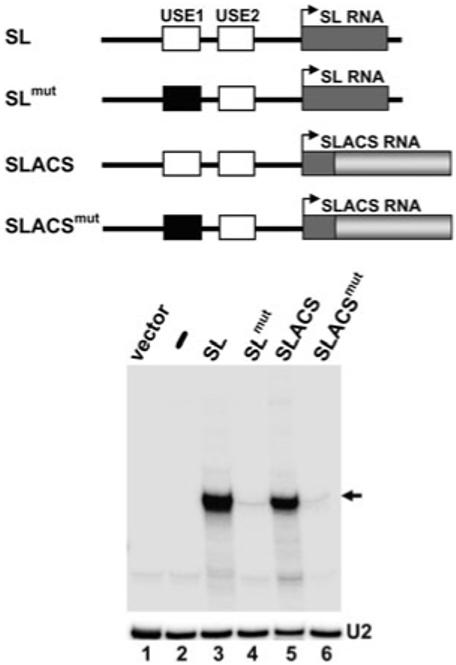
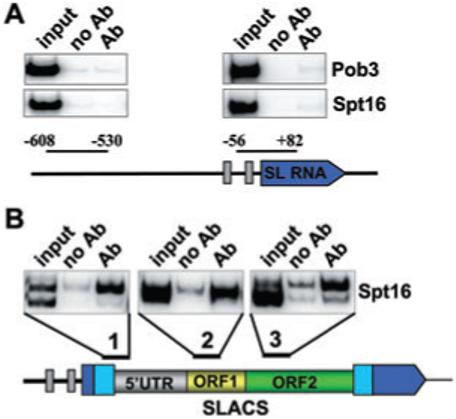
The Trypanosoma brucei genome is colonized by the site-specific non-LTR retrotransposon SLACS, or spliced leader-associated conserved sequence, which integrates exclusively into the spliced leader (SL) RNA genes. Although there is evidence that the RNA interference (RNAi) machinery regulates SLACS transcript levels, we do not know whether RNAi deficiency affects the genomic stability of SLACS, nor do we understand the mechanism of SLACS transcription. Here, we report that prolonged culturing of RNAi-deficient T. brucei cells, but not wild-type cells, results in genomic rearrangements of SLACS. Furthermore, two populations of SLACS transcripts persist in RNAi-deficient cells: a full-length transcript of approximately 7 kb and a heterogeneous population of small SLACS transcripts ranging in size from 450 to 550 nt. We provide evidence that SLACS transcription initiates at the +1 of the interrupted SL RNA gene and proceeds into the 5' UTR and open reading frame 1 (ORF1). This transcription is carried out by an RNA polymerase with alpha-amanitin sensitivity reminiscent of SL RNA synthesis and is dependent on the SL RNA promoter. Additionally, we show that both sense and antisense small SLACS transcripts originate from ORF1 and that they are associated with proteins in vivo. We speculate that the small SLACS transcripts serve as substrates for the production of siRNAs to regulate SLACS expression.
Figures
References
-
- Aravin AA, Naumova NM, Tulin AV, Vagin VV, Rozovsky YM, Gvozdev VA. Double-stranded RNA-mediated silencing of genomic tandem repeats and transposable elements in the D. melanogaster germline. Curr Biol. 2001;11:1017–1027. - PubMed
-
- Axtell MJ, Jan C, Rajagopalan R, Bartel DP. A two-hit trigger for siRNA biogenesis in plants. Cell. 2006;127:565–577. - PubMed
-
- Baulcombe DC. Molecular biology. Amplified silencing. Science. 2007;315:199–200. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
