

doi: 10.1093/nar/gkm1063.
Epub 2007 Dec 10.
Quantitative telomeric overhang determination using a double-strand specific nuclease
Affiliations
- PMID: 18073199
- PMCID: PMC2241892
- DOI: 10.1093/nar/gkm1063
Item in Clipboard
Quantitative telomeric overhang determination using a double-strand specific nuclease
Nucleic Acids Res.
2008 Feb.
Abstract
Telomeres terminate in 3' overhangs that function in end protection and the formation of t-loops. Determining the steps and factors involved in overhang processing is compromised by the inability to easily and accurately determine overhang size in the presence of many kilobases of double-stranded telomeric DNA. We here describe the use of a double-strand specific nuclease (DSN) that entirely digests double-stranded DNA including telomeres, leaving the overhangs intact so that they can be measured.
Figures
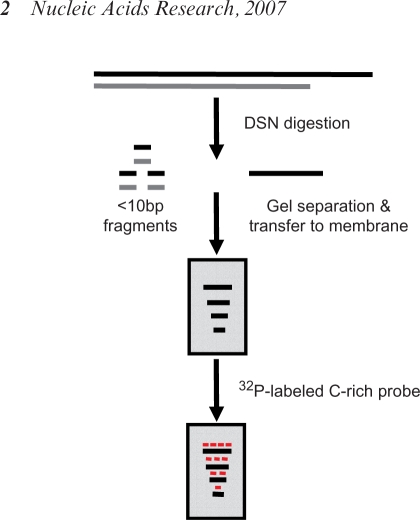
Strategy of the DSN base overhang assay. Complete and specific digestion of all double-stranded DNA should leave the single-stranded TTAGGG sequences present in the telomeric 3′ overhangs available for analysis on denaturing gels.
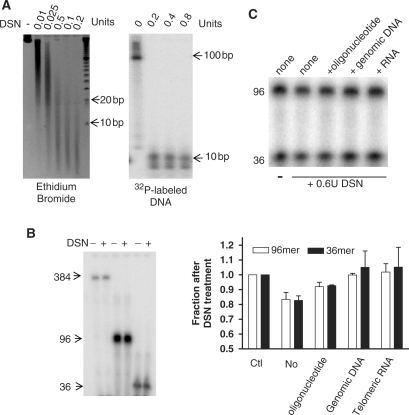
Characterization of DSN in telomere overhang assays. (A) DSN digests ds telomeres to <10 bp. Left: Five microgram of Hela genomic DNA was digested with DSN and analyzed on 6% polyacrylamide gel with ethidium bromide staining for total DNA. Right: One nanogram of ds telomeric DNA (96 bp, continuously 32P-labeled by primer extension) was added to 5 μg genomic DNA as above. (B) Single-stranded telomeric DNA is resistant to DSN digestion. Approximately two nanomolar of 32P-end-labeled oligonucleotides were digested with 0.2 U of DSN at 37°C for 2 h. (C) Minor ss activity of DSN is eliminated by excess competitor. Top: End-labeled telomeric oligonucleotides (0.5 μM each of 16 and 6 repeats in a 20 μl reaction) were digested with 0.6 U of DSN at 37°C for 1 h. Two microgram of different competitors (a 33-mer random oligonucleotide, genomic DNA from HT1080 cells or a UUAGGGUUAGGGUUAGGGUU RNA) were added. Bottom: The fraction remaining after DSN digestion in the presence of various competitors was quantitated in two experiments, ±SD.
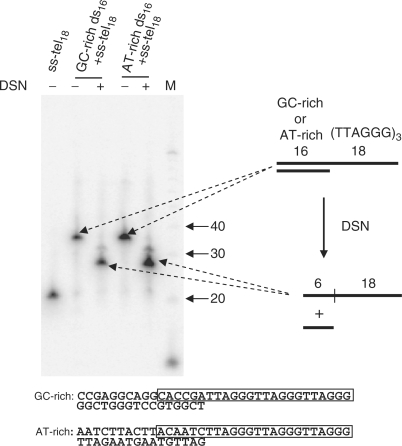
Five to six ds junctional nucleotides are resistant to DSN treatment. Here, 34 nt oligonucleotides containing either 16 bp of AT-rich or GC-rich sequence followed by 18 nt of ss telomeric sequence were incubated with or without DSN. After incubation, the DNA was loaded onto a 10% polyacrylamide gel with 8 M urea and DNA was then transferred to a membrane and hybridized with a hot C-rich telomeric probe. The sequences of the annealed oligonucleotides are shown, and the regions surviving DSN digestion are boxed. Five to six nucleotides of ds DNA remain undigested regardless of the specific ds–ss junctional sequence.
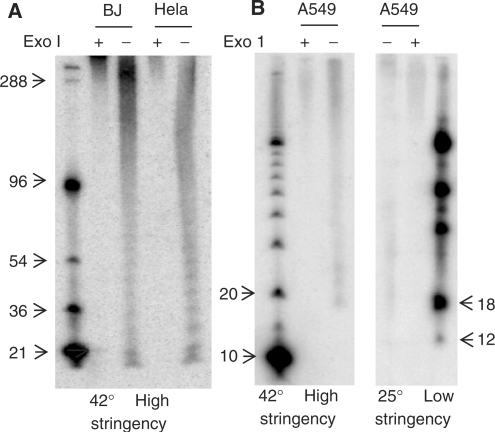
DSN detects human telomeres as short as 12 nt. (A) Representative DSN overhang assays on BJ fibroblast and Hela adenocarcinoma DNA. Results are quantitated in Table 1. ExoI pretreatment removes the 3′ overhang. (B) A sharp signal loss below 18 nt under normal high stringency hybridization condition is also seen when the membrane was hybridized at low stringency. The signal at 18 nt corresponds to an overhang length of 12 nt.
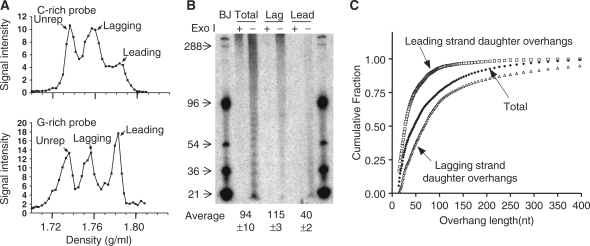
Human leading strand daughter telomeres have very short overhangs. (A) Separation of telomeric leading and lagging strands from BJ cells grown in BrdU for 48 h using CsCl gradients (slot blot results). The signal from leading strand daughter telomeres appeared to be less than that from lagging telomeres only when a C-rich probe was used. (B) Overhangs at lagging and leading strand daughter telomeres in normal fibroblasts. Averages are mean ± SD, n = 2. (C) Distribution of overhang sizes. The cumulative fraction of overhangs shorter than a given size is plotted after quantitating the signals in (B) to illustrate the distribution of lengths.
References
-
- Blackburn EH. Switching and signaling at the telomere. Cell. 2001;106:661–673. - PubMed
-
- de Lange T. Protection of mammalian telomeres. Oncogene. 2002;21:532–540. - PubMed
-
- Tahara H, Kusunoki M, Yamanaka Y, Matsumura S, Ide T. G-tail telomere HPA: simple measurement of human single-stranded telomeric overhangs. Nat. Methods. 2005;2:829–831. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
