

Investigating the genetic circuitry of mastermind in Drosophila, a notch signal effector
- PMID: 18073442
- PMCID: PMC2219471
- DOI: 10.1534/genetics.107.080994
Investigating the genetic circuitry of mastermind in Drosophila, a notch signal effector
Abstract
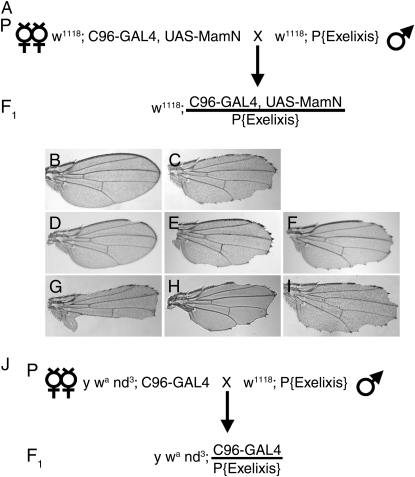
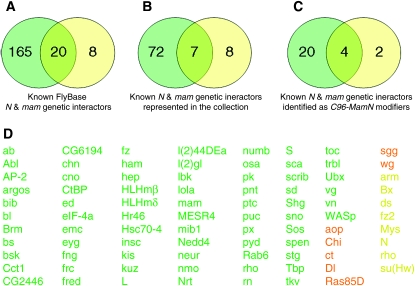
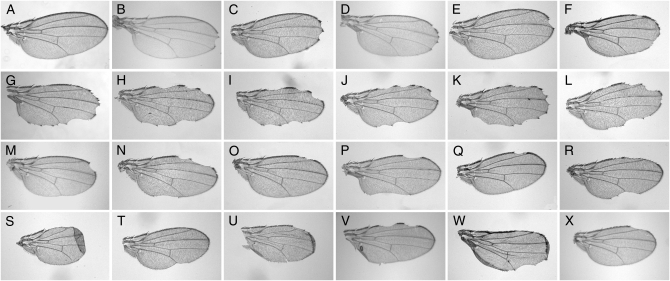
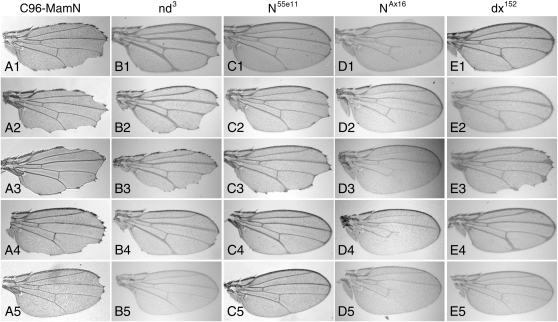
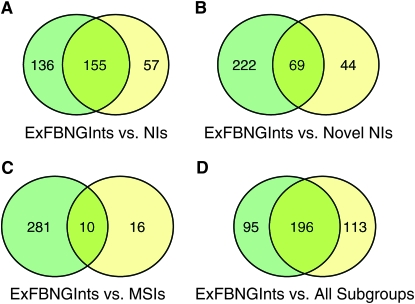
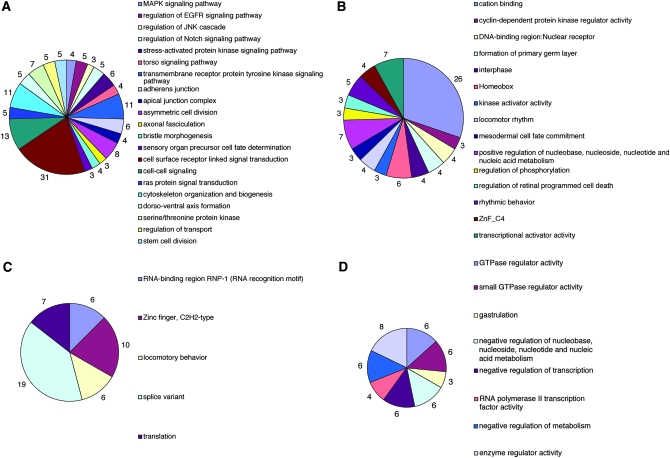
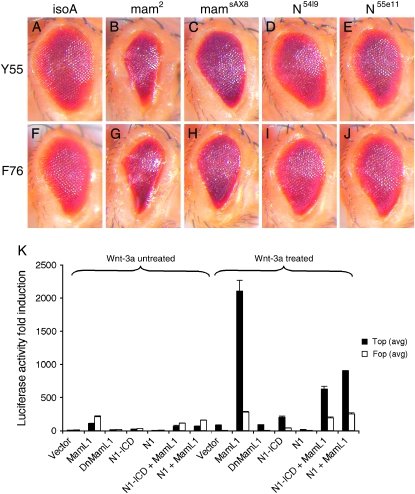
Notch signaling regulates multiple developmental processes and is implicated in various human diseases. Through use of the Notch transcriptional co-activator mastermind, we conducted a screen for Notch signal modifiers using the Exelixis collection of insertional mutations, which affects approximately 50% of the Drosophila genome, recovering 160 genes never before associated with Notch, extending the previous roster of genes that interact functionally with the Notch pathway and mastermind. As the molecular identity for most recovered genes is known, gene ontology (GO) analysis was applied, grouping genes according to functional classifications. We identify novel Notch-associated GO categories, uncover nodes of integration between Notch and other signaling pathways, and unveil groups of modifiers that suggest the existence of Notch-independent mastermind functions, including a conserved ability to regulate Wnt signaling.
Figures

Similar articles
-
Characterization of a new mastermind allele identified from somatic mosaic screen.Cells Dev. 2021 Mar;165:203664. doi: 10.1016/j.cdev.2021.203664. Epub 2021 Feb 4. Cells Dev. 2021. PMID: 33993981
-
MAML1, a human homologue of Drosophila mastermind, is a transcriptional co-activator for NOTCH receptors.Nat Genet. 2000 Dec;26(4):484-9. doi: 10.1038/82644. Nat Genet. 2000. PMID: 11101851
-
Dynamic modes of Notch transcription hubs conferring memory and stochastic activation revealed by live imaging the co-activator Mastermind.Elife. 2024 May 10;12:RP92083. doi: 10.7554/eLife.92083. Elife. 2024. PMID: 38727722 Free PMC article.
-
Notch signalling in the nucleus: roles of Mastermind-like (MAML) transcriptional coactivators.J Biochem. 2016 Mar;159(3):287-94. doi: 10.1093/jb/mvv123. Epub 2015 Dec 28. J Biochem. 2016. PMID: 26711237 Review.
-
Notch signaling: simplicity in design, versatility in function.Development. 2011 Sep;138(17):3593-612. doi: 10.1242/dev.063610. Development. 2011. PMID: 21828089 Review.
Cited by
-
A targeted genetic modifier screen links the SWI2/SNF2 protein domino to growth and autophagy genes in Drosophila melanogaster.G3 (Bethesda). 2013 May 20;3(5):815-25. doi: 10.1534/g3.112.005496. G3 (Bethesda). 2013. PMID: 23550128 Free PMC article.
-
Synergy between the ESCRT-III complex and Deltex defines a ligand-independent Notch signal.J Cell Biol. 2011 Dec 12;195(6):1005-15. doi: 10.1083/jcb.201104146. J Cell Biol. 2011. PMID: 22162134 Free PMC article.
-
Nemo-like kinase suppresses Notch signalling by interfering with formation of the Notch active transcriptional complex.Nat Cell Biol. 2010 Mar;12(3):278-85. doi: 10.1038/ncb2028. Epub 2010 Jan 31. Nat Cell Biol. 2010. PMID: 20118921
-
High Throughput Sequencing Identifies Misregulated Genes in the Drosophila Polypyrimidine Tract-Binding Protein (hephaestus) Mutant Defective in Spermatogenesis.PLoS One. 2016 Mar 4;11(3):e0150768. doi: 10.1371/journal.pone.0150768. eCollection 2016. PLoS One. 2016. PMID: 26942929 Free PMC article.
-
A polycistronic locus encodes both a tumor-suppressive microRNA and a long non-coding RNA regulating testis morphology in Drosophila.bioRxiv [Preprint]. 2025 May 16:2024.10.10.617551. doi: 10.1101/2024.10.10.617551. bioRxiv. 2025. PMID: 39416153 Free PMC article. Preprint.
References
-
- Alves-Guerra, M. C., C. Ronchini and A. J. Capobianco, 2007. Mastermind-like 1 is a specific coactivator of β-catenin transcription activation and is essential for colon carcinoma cell survival. Cancer Res. 67: 8690–8698. - PubMed
-
- Armstrong, J. A., A. S. Sperling, R. Deuring, L. Manning, S. L. Moseley et al., 2005. Genetic screens for enhancers of brahma reveal functional interactions between the BRM chromatin-remodeling complex and the delta-notch signal transduction pathway in Drosophila. Genetics 170: 1761–1774. - PMC - PubMed
-
- Arn, E. A., B. J. Cha, W. E. Theurkauf and P. M. Macdonald, 2003. Recognition of a bicoid mRNA localization signal by a protein complex containing Swallow, Nod, and RNA binding proteins. Dev. Cell 4: 41–51. - PubMed
-
- Artavanis-Tsakonas, S., 2004. Accessing the Exelixis collection. Nat. Genet. 36: 207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
