

Evolution of allostery in the cyclic nucleotide binding module
- PMID: 18076763
- PMCID: PMC2246266
- DOI: 10.1186/gb-2007-8-12-r264
Evolution of allostery in the cyclic nucleotide binding module
Abstract
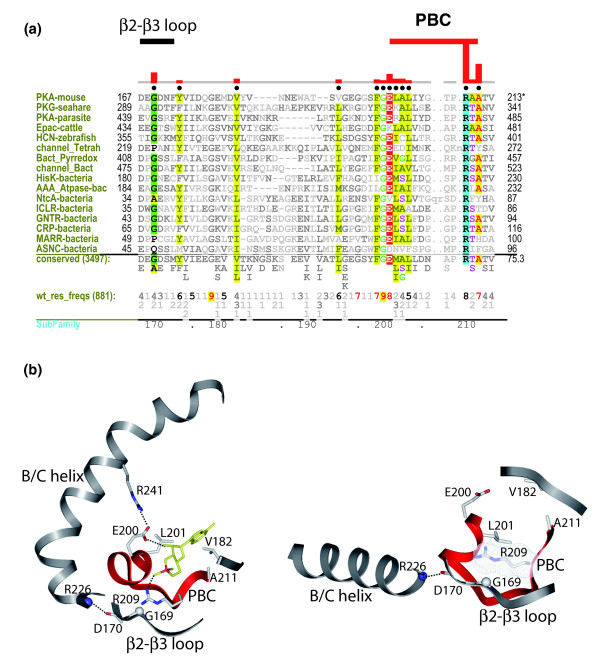
Background: The cyclic nucleotide binding (CNB) domain regulates signaling pathways in both eukaryotes and prokaryotes. In this study, we analyze the evolutionary information embedded in genomic sequences to explore the diversity of signaling through the CNB domain and also how the CNB domain elicits a cellular response upon binding to cAMP.
Results: Identification and classification of CNB domains in Global Ocean Sampling and other protein sequences reveals that they typically are fused to a wide variety of functional domains. CNB domains have undergone major sequence variation during evolution. In particular, the sequence motif that anchors the cAMP phosphate (termed the PBC motif) is strikingly different in some families. This variation may contribute to ligand specificity inasmuch as members of the prokaryotic cooA family, for example, harbor a CNB domain that contains a non-canonical PBC motif and that binds a heme ligand in the cAMP binding pocket. Statistical comparison of the functional constraints imposed on the canonical and non-canonical PBC containing sequences reveals that a key arginine, which coordinates with the cAMP phosphate, has co-evolved with a glycine in a distal beta2-beta3 loop that allosterically couples cAMP binding to distal regulatory sites.
Conclusion: Our analysis suggests that CNB domains have evolved as a scaffold to sense a wide variety of second messenger signals. Based on sequence, structural and biochemical data, we propose a mechanism for allosteric regulation by CNB domains.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
