

Reduced mitochondrial SOD displays mortality characteristics reminiscent of natural aging
- PMID: 18078670
- PMCID: PMC2675272
- DOI: 10.1016/j.mad.2007.10.013
Reduced mitochondrial SOD displays mortality characteristics reminiscent of natural aging
Abstract
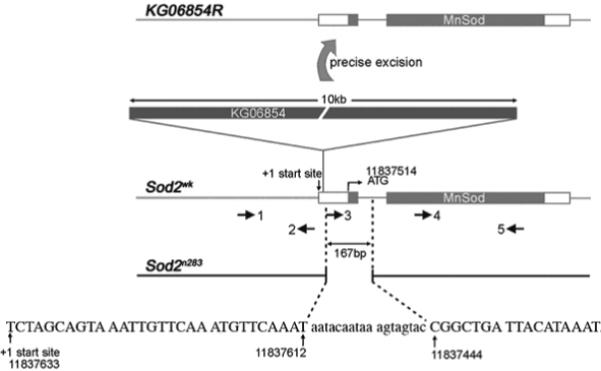
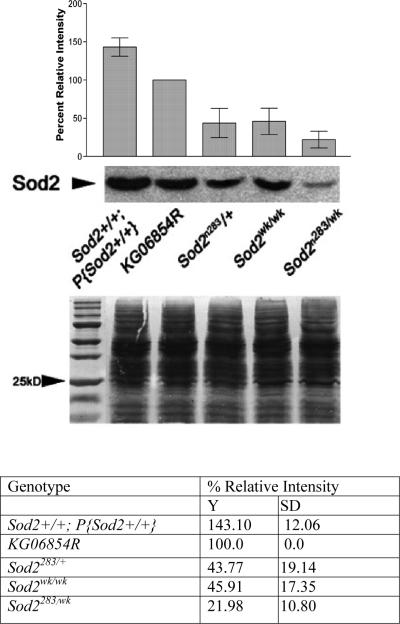
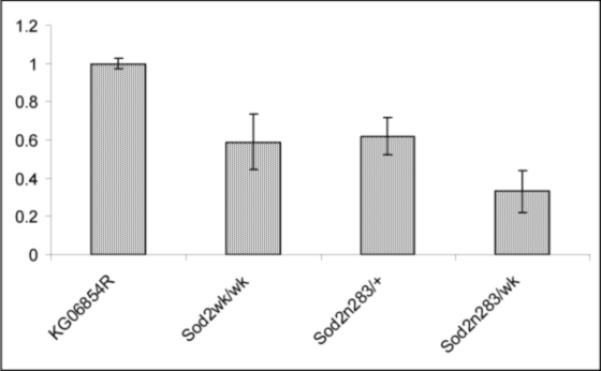
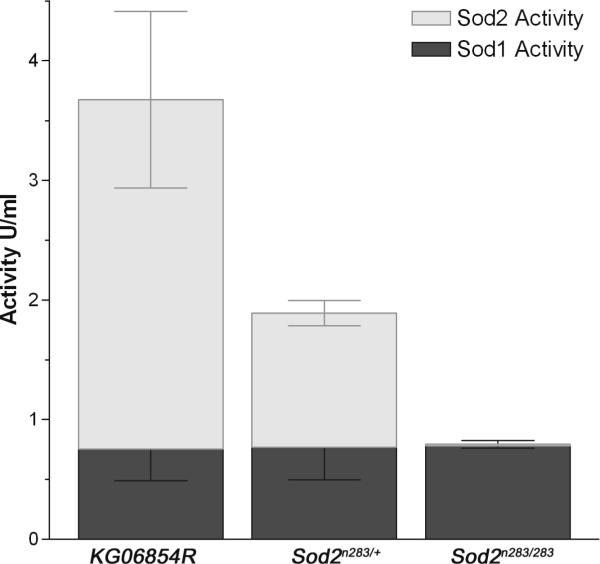
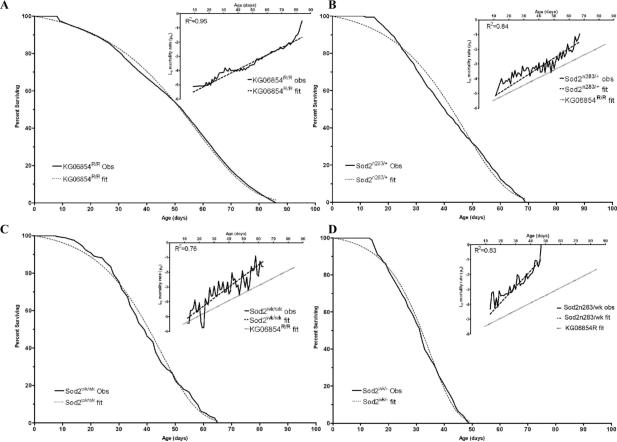
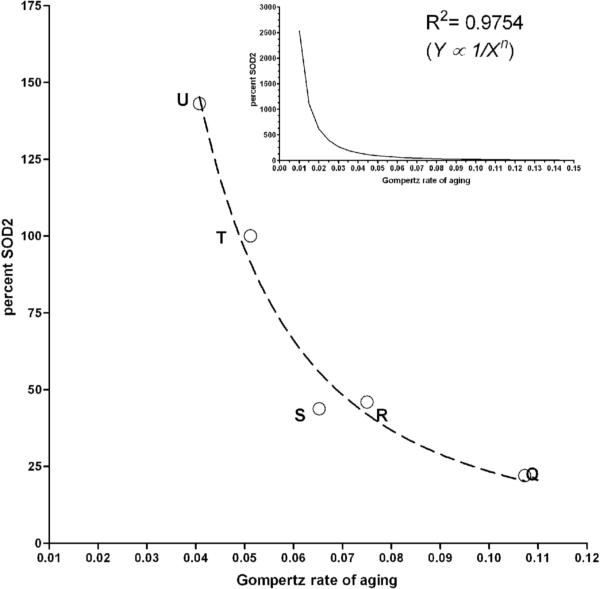
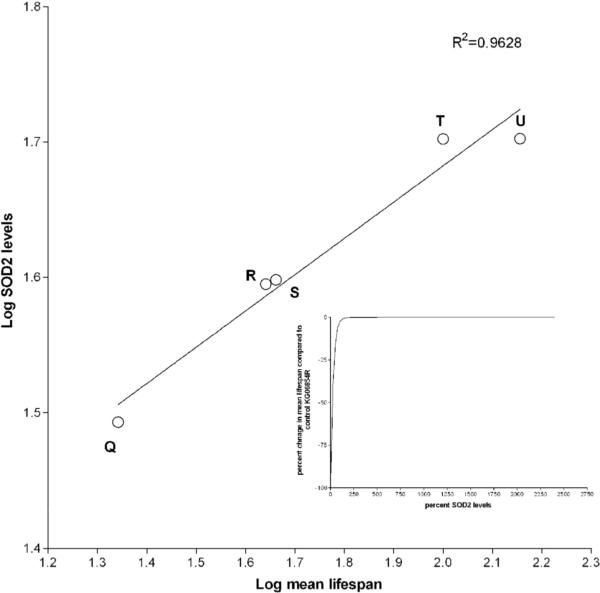
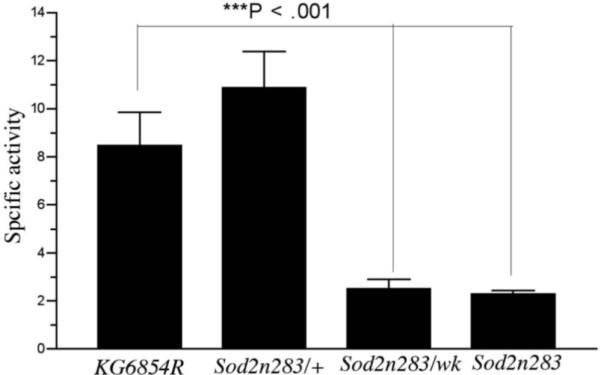
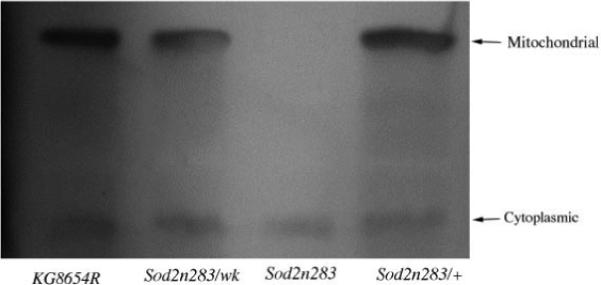
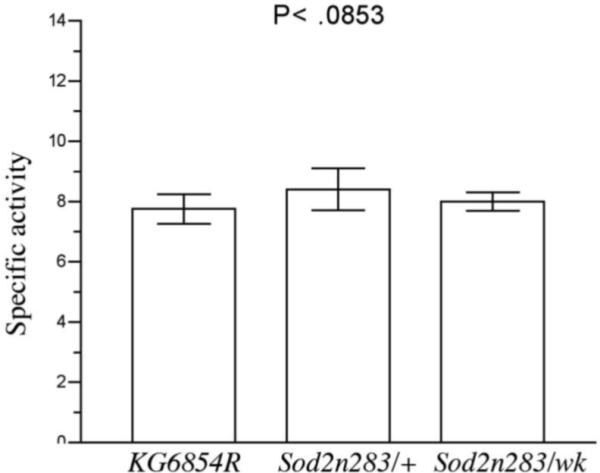
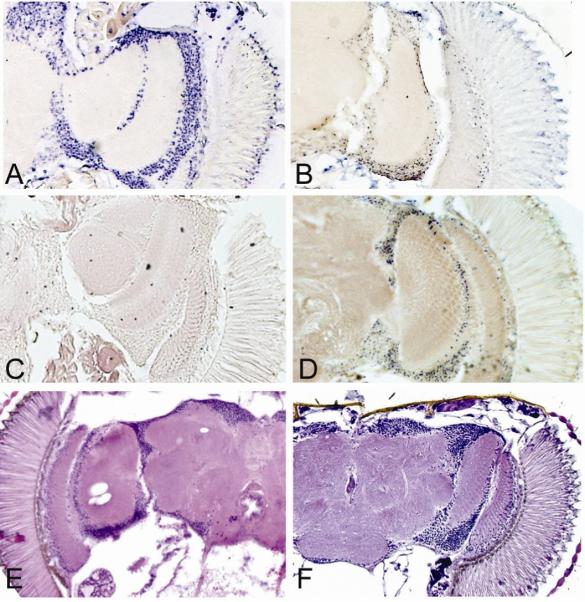
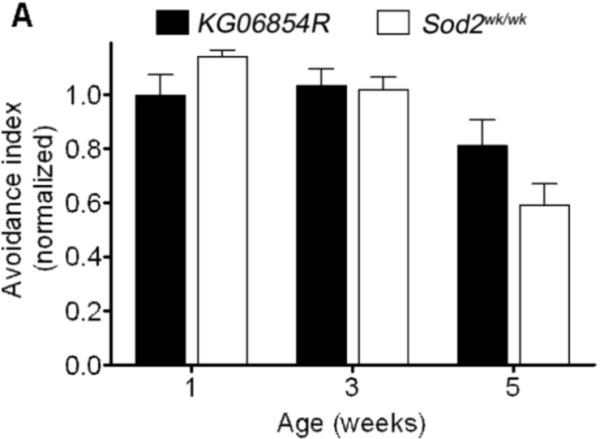
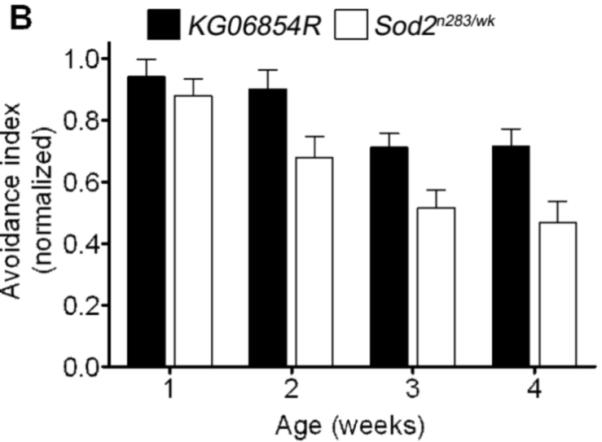
Manganese superoxide dismutase (MnSOD or SOD2) is a key mitochondrial enzymatic antioxidant. Arguably the most striking phenotype associated with complete loss of SOD2 in flies and mice is shortened life span. To further explore the role of SOD2 in protecting animals from aging and age-associated pathology, we generated a unique collection of Drosophila mutants that progressively reduce SOD2 expression and function. Mitochondrial aconitase activity was substantially reduced in the Sod2 mutants, suggesting that SOD2 normally ensures the functional capacity of mitochondria. Flies with severe reductions in SOD2 expression exhibited accelerated senescence of olfactory behavior as well as precocious neurodegeneration and DNA strand breakage in neurons. Furthermore, life span was progressively shortened and age-dependent mortality was increased in conjunction with reduced SOD2 expression, while initial mortality and developmental viability were unaffected. Interestingly, life span and age-dependent mortality varied exponentially with SOD2 activity, indicating that there might normally be a surplus of this enzyme for protecting animals from premature death. Our data support a model in which disruption of the protective effects of SOD2 on mitochondria manifests as profound changes in behavioral and demographic aging as well as exacerbated age-related pathology in the nervous system.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
