

Cell-cycle regulation of cohesin stability along fission yeast chromosomes
- PMID: 18079700
- PMCID: PMC2206136
- DOI: 10.1038/sj.emboj.7601955
Cell-cycle regulation of cohesin stability along fission yeast chromosomes
Abstract
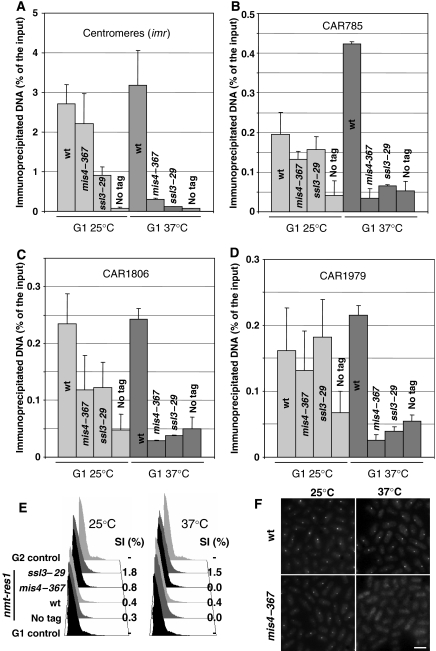
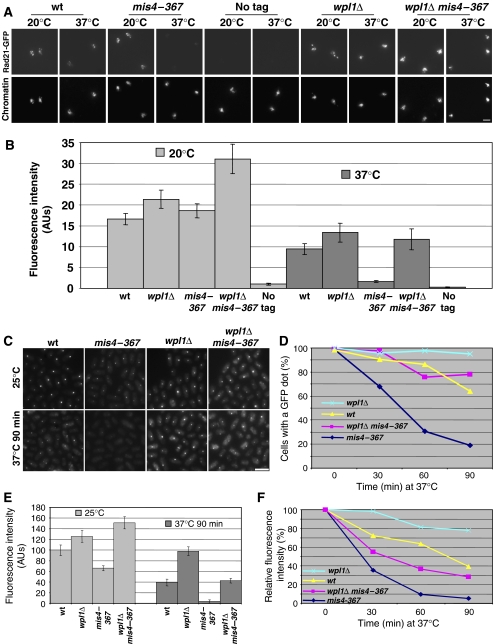
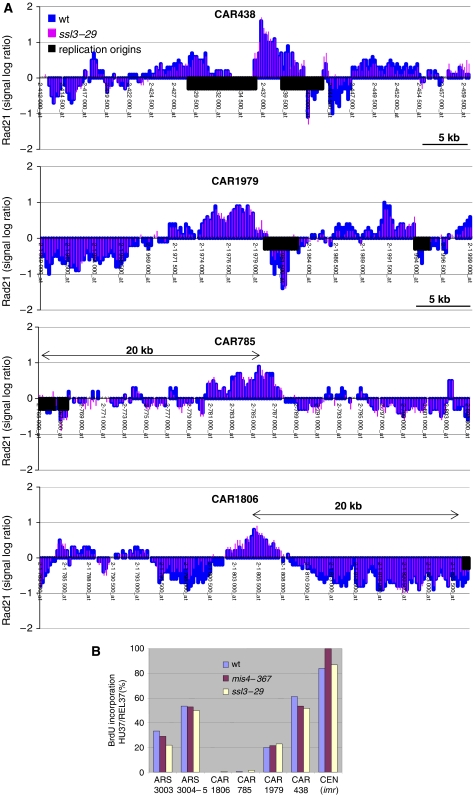
Sister chromatid cohesion is mediated by cohesin, but the process of cohesion establishment during S-phase is still enigmatic. In mammalian cells, cohesin binding to chromatin is dynamic in G1, but becomes stabilized during S-phase. Whether the regulation of cohesin stability is integral to the process of cohesion establishment is unknown. Here, we provide evidence that fission yeast cohesin also displays dynamic behavior. Cohesin association with G1 chromosomes requires continued activity of the cohesin loader Mis4/Ssl3, suggesting that repeated loading cycles maintain cohesin binding. Cohesin instability in G1 depends on wpl1, the fission yeast ortholog of mammalian Wapl, suggestive of a conserved mechanism that controls cohesin stability on chromosomes. wpl1 is nonessential, indicating that a change in wpl1-dependent cohesin dynamics is dispensable for cohesion establishment. Instead, we find that cohesin stability increases at the time of S-phase in a reaction that can be uncoupled from DNA replication. Hence, cohesin stabilization might be a pre-requisite for cohesion establishment rather than its consequence.
Figures



References
-
- Arumugam P, Gruber S, Tanaka K, Haering CH, Mechtler K, Nasmyth K (2003) ATP hydrolysis is required for cohesin's association with chromosomes. Curr Biol 13: 1941–1953 - PubMed
-
- Bahler J, Wu JQ, Longtine MS, Shah NG, McKenzie A III, Steever AB, Wach A, Philippsen P, Pringle JR (1998) Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast 14: 943–951 - PubMed
-
- Bernard P, Drogat J, Maure JF, Dheur S, Vaur S, Genier S, Javerzat JP (2006) A screen for cohesion mutants uncovers Ssl3, the fission yeast counterpart of the cohesin loading factor Scc4. Curr Biol 16: 875–881 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
