

Sequential recruitment of the repair factors during NER: the role of XPG in initiating the resynthesis step
- PMID: 18079701
- PMCID: PMC2206130
- DOI: 10.1038/sj.emboj.7601948
Sequential recruitment of the repair factors during NER: the role of XPG in initiating the resynthesis step
Abstract
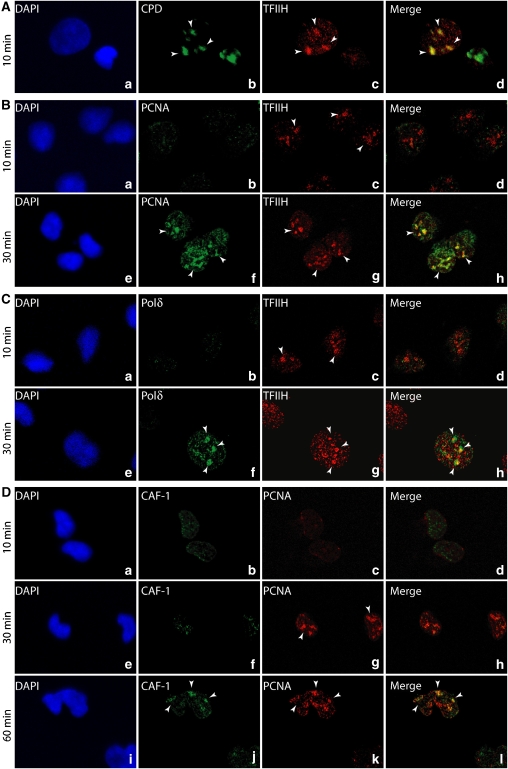
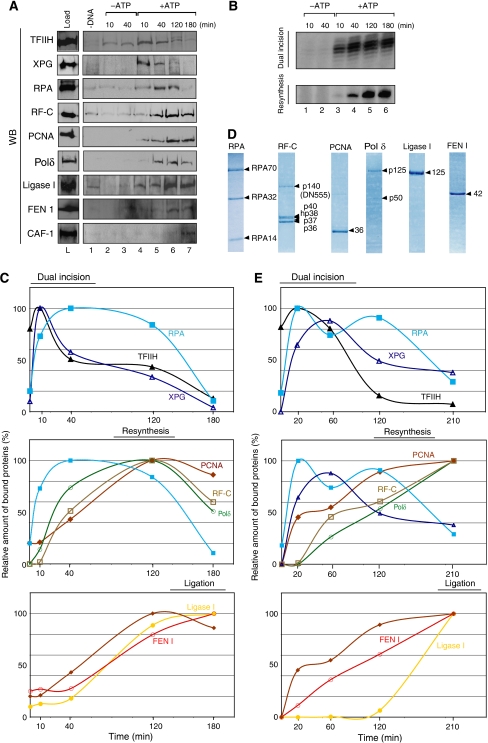
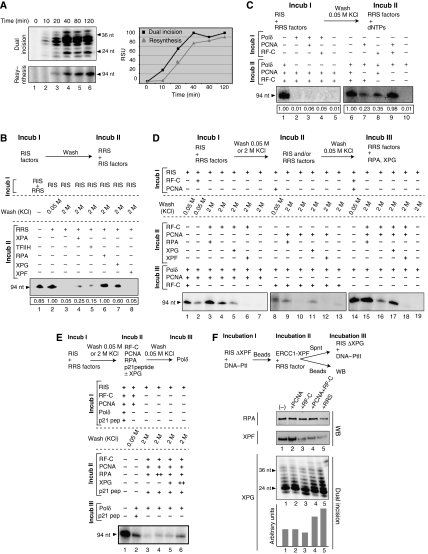
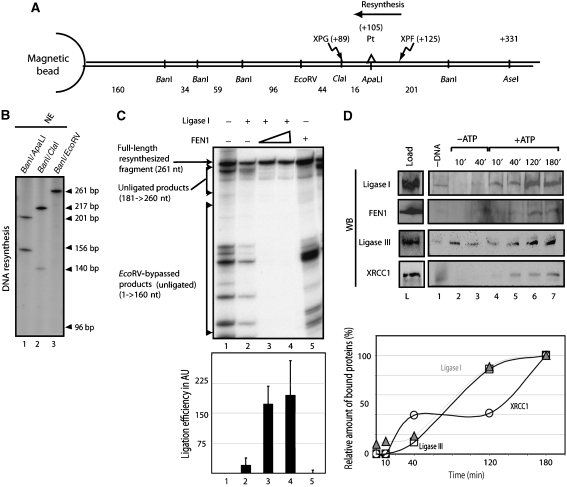
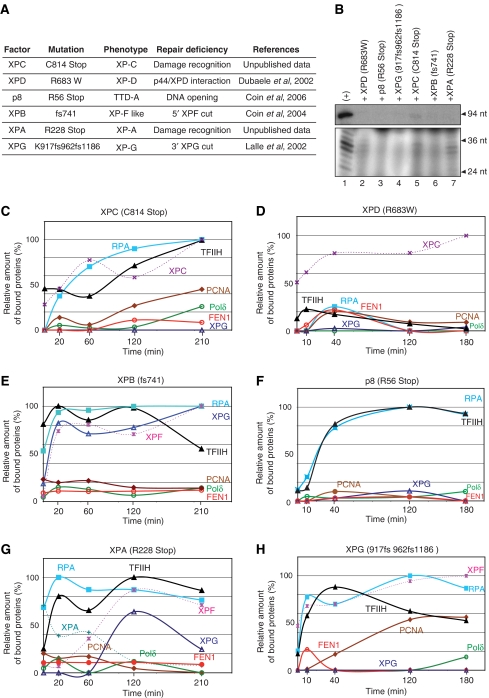
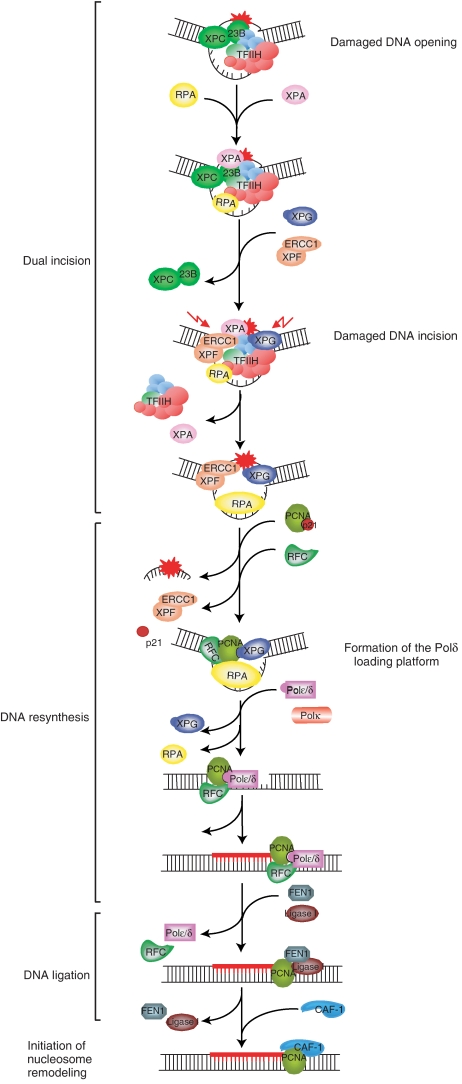
To address the biochemical mechanisms underlying the coordination between the various proteins required for nucleotide excision repair (NER), we employed the immobilized template system. Using either wild-type or mutated recombinant proteins, we identified the factors involved in the NER process and showed the sequential comings and goings of these factors to the immobilized damaged DNA. Firstly, we found that PCNA and RF-C arrival requires XPF 5' incision. Moreover, the positioning of RF-C is facilitated by RPA and induces XPF release. Concomitantly, XPG leads to PCNA recruitment and stabilization. Our data strongly suggest that this interaction with XPG protects PCNA and Pol delta from the effect of inhibitors such as p21. XPG and RPA are released as soon as Pol delta is recruited by the RF-C/PCNA complex. Finally, a ligation system composed of FEN1 and Ligase I can be recruited to fully restore the DNA. In addition, using XP or trichothiodystrophy patient-derived cell extracts, we were able to diagnose the biochemical defect that may prove to be important for therapeutic purposes.
Figures
References
-
- Aboussekhra A, Biggerstaff M, Shivji MK, Vilpo JA, Moncollin V, Podust VN, Protic M, Hubscher U, Egly JM, Wood RD (1995) Mammalian DNA nucleotide excision repair reconstituted with purified protein components. Cell 80: 859–868 - PubMed
-
- Araujo SJ, Wood RD (1999) Protein complexes in nucleotide excision repair. Mutat Res 435: 23–33 - PubMed
-
- Ayyagari R, Gomes XV, Gordenin DA, Burgers PM (2003) Okazaki fragment maturation in yeast. I. Distribution of functions between FEN1 AND DNA2. J Biol Chem 278: 1618–1625 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
