

S2 deletion variants of human PRL receptors demonstrate that extracellular domain conformation can alter conformation of the intracellular signaling domain
- PMID: 18081308
- PMCID: PMC2408824
- DOI: 10.1021/bi7013882
S2 deletion variants of human PRL receptors demonstrate that extracellular domain conformation can alter conformation of the intracellular signaling domain
Abstract
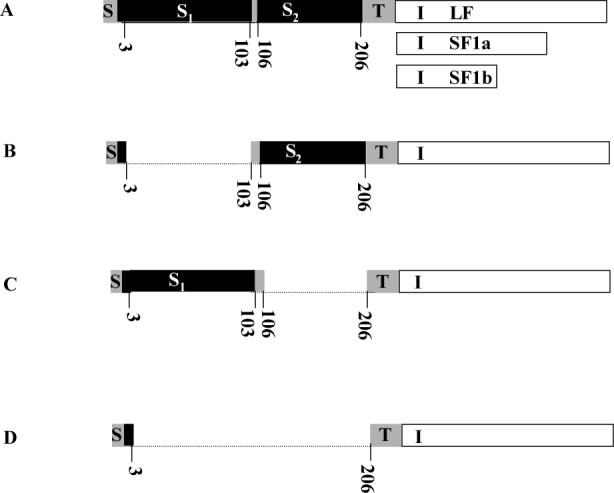
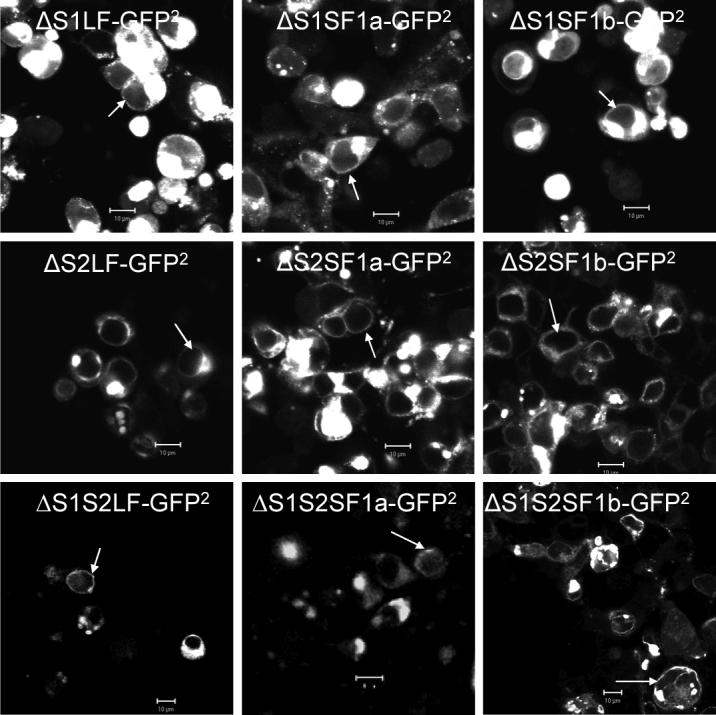
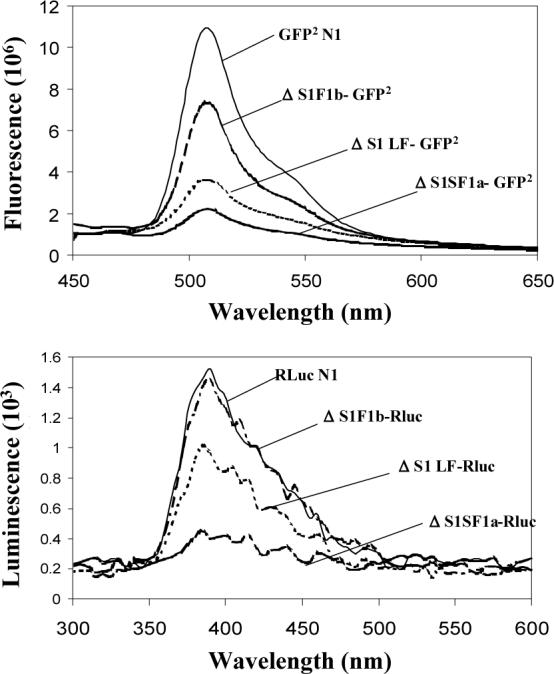
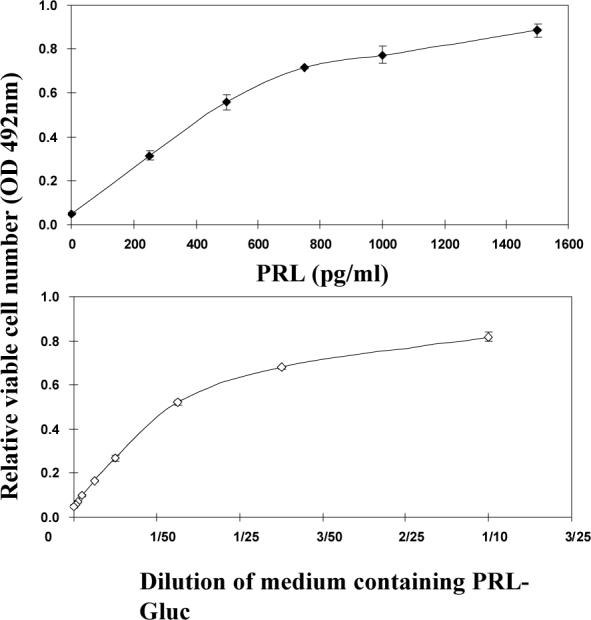
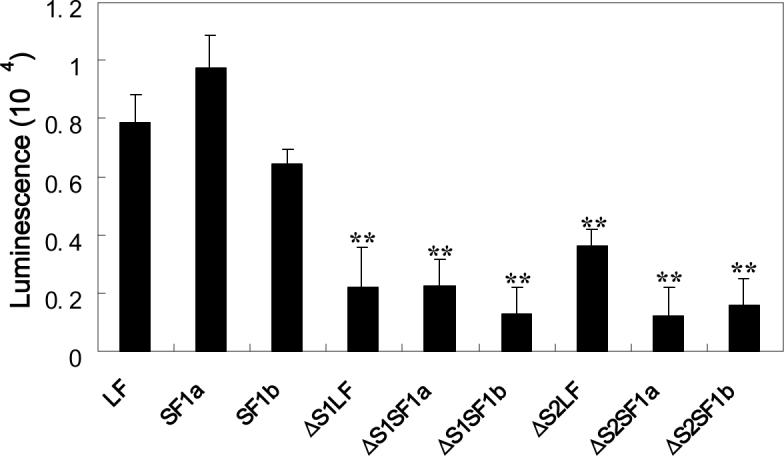
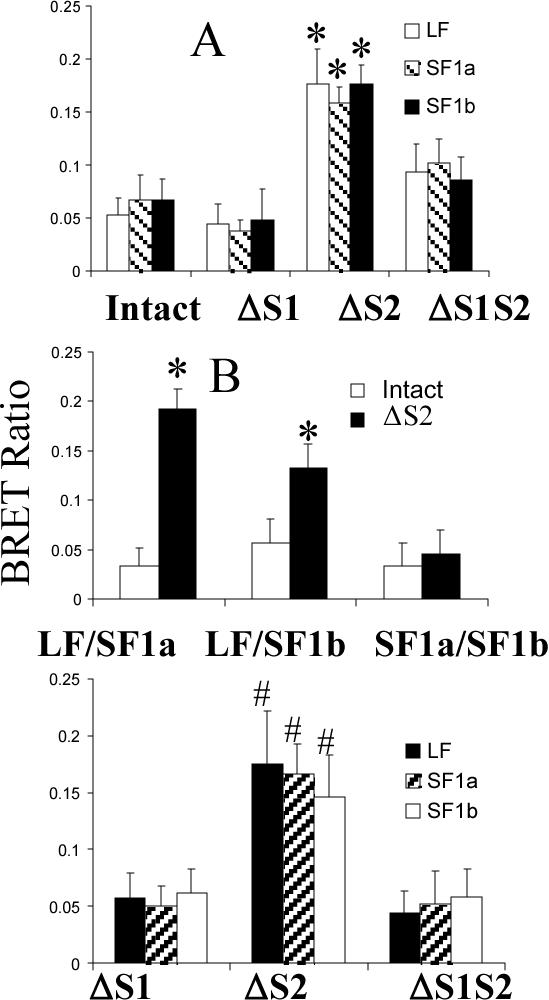
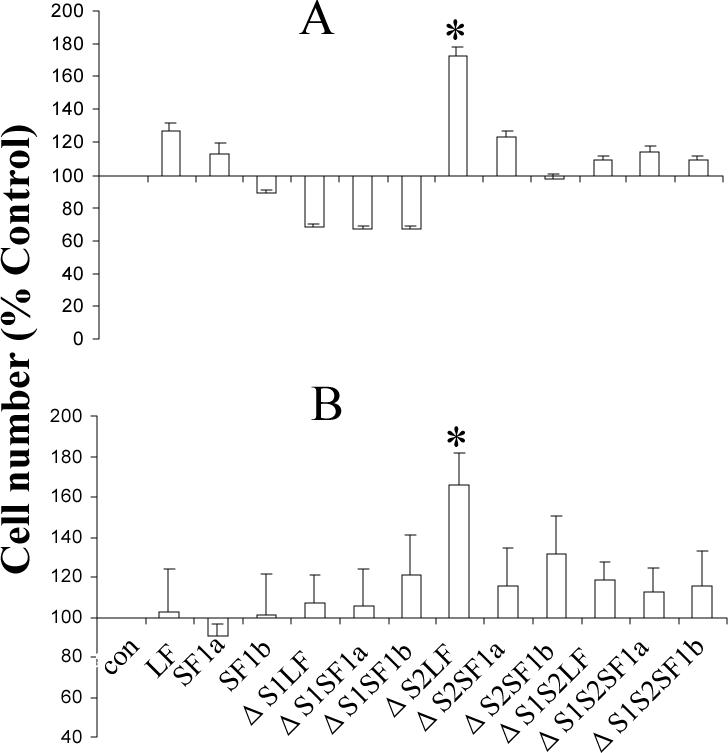
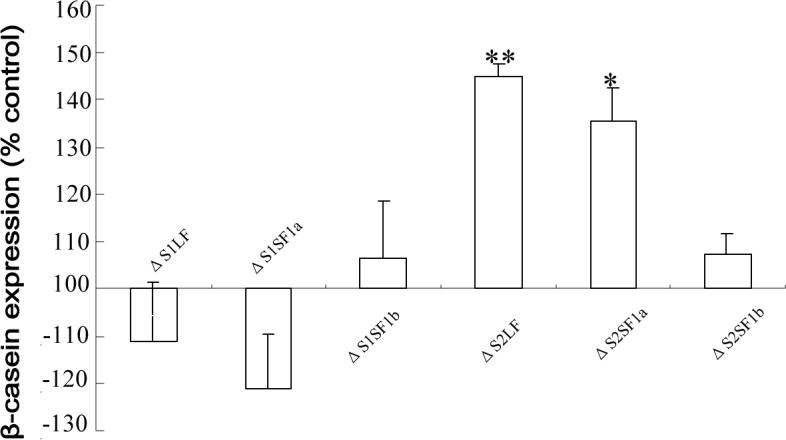
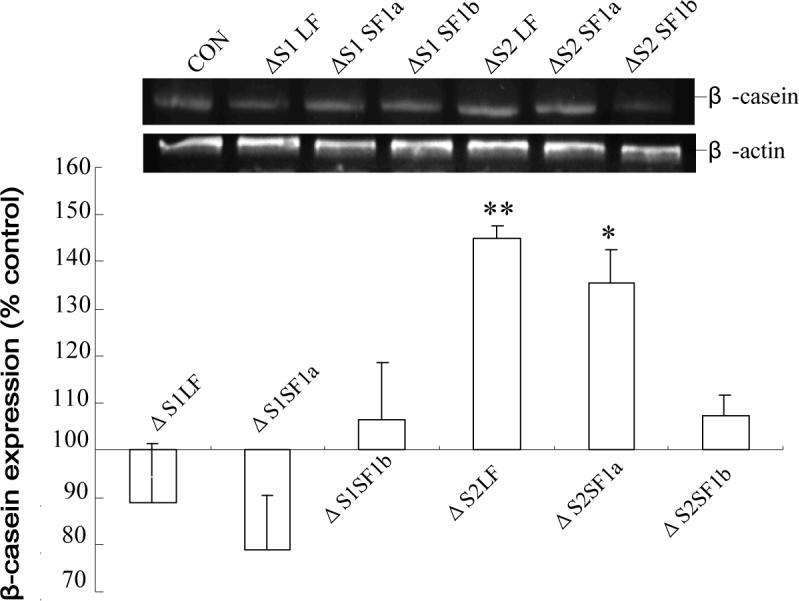
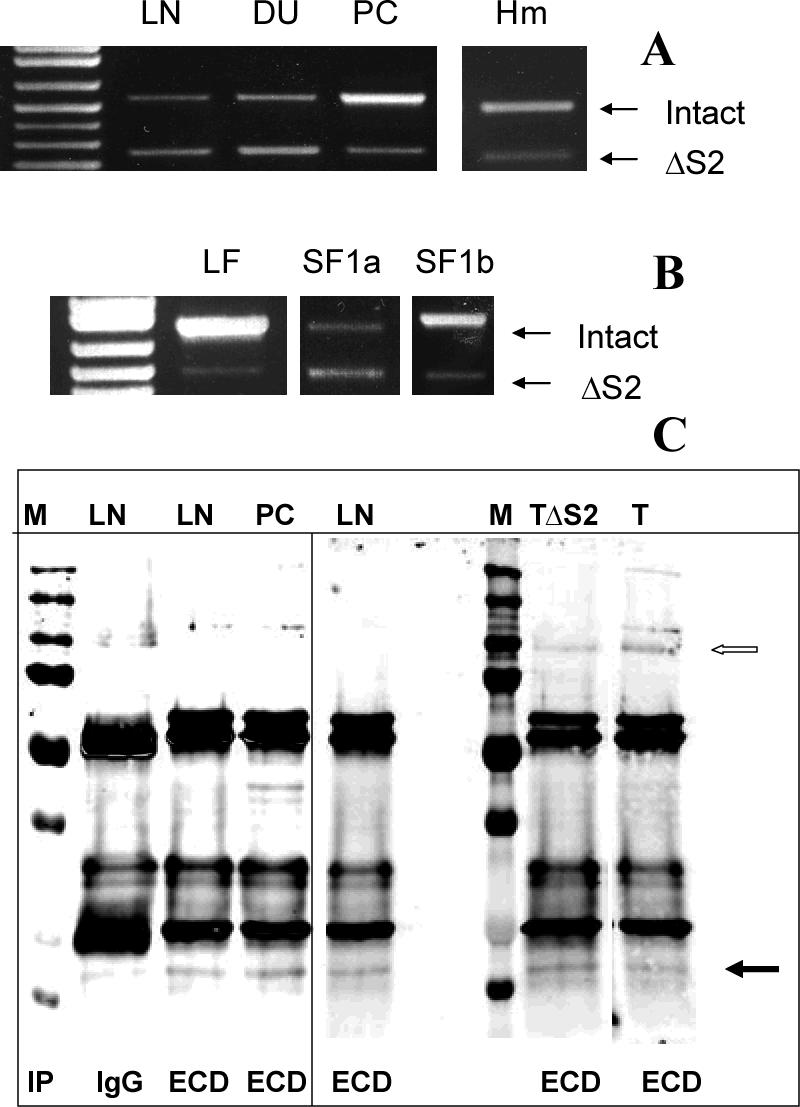
Using spacers between the C-termini of the long (LF) or short (SF) human prolactin receptors and luciferase/GFP such that bioluminescence resonance energy transfer (BRET) occurred minimally in intact versions of these receptors in the absence of ligand, we have monitored the BRET signal after deletion of regions of the extracellular domain (ECD). Deletion of S2 produced ligand-independent BRET for only those pairings normally occurring in the presence of ligand with the intact receptor. Deletion of the similarly sized S1, or S1 plus S2, produced no ligand-independent or -dependent BRET. When deleted receptors were transfected into human breast (T47D) or prostate (DU145) cancer cells incubated in the absence of added prolactin (PRL) and presence of anti-PRL, expression of the DeltaS2LF resulted in increased cell number, whereas expression of the intact receptor did not. When endogenous beta-casein expression was examined in T47D cells, the DeltaS2LF and DeltaS2F1a both showed ligand-independent activation of transcription, again not duplicated by the intact receptor. Paired with evidence in the literature for predimerization of PRLRs, these results demonstrate that altered ECD conformation, and not just a change in bulk, produces altered conformation of the intracellular signaling region of the receptors, supporting the concept that ligand binding to the ECD of intact predimerized receptors could initiate signaling. In addition, the current work supports a dual proliferative and differentiative role for the LF receptor, but only a differentiative role for the SF1a receptor. Naturally occurring DeltaS2 PRL receptors (PRLR) were also found in normal and cancerous human cells. This additionally suggests a heretofore unappreciated ligand-independent role for PRLRs.
Figures
Similar articles
-
Unmodified prolactin (PRL) and S179D PRL-initiated bioluminescence resonance energy transfer between homo- and hetero-pairs of long and short human PRL receptors in living human cells.Mol Endocrinol. 2005 May;19(5):1291-303. doi: 10.1210/me.2004-0304. Epub 2005 Feb 3. Mol Endocrinol. 2005. PMID: 15695371
-
Ligand-independent dimerization of the human prolactin receptor isoforms: functional implications.Mol Endocrinol. 2006 Nov;20(11):2734-46. doi: 10.1210/me.2006-0114. Epub 2006 Jul 13. Mol Endocrinol. 2006. PMID: 16840534
-
Functional characterization of the intermediate isoform of the human prolactin receptor.J Biol Chem. 1999 Dec 10;274(50):35461-8. doi: 10.1074/jbc.274.50.35461. J Biol Chem. 1999. PMID: 10585417
-
Prolactin (PRL) and its receptor: actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice.Endocr Rev. 1998 Jun;19(3):225-68. doi: 10.1210/edrv.19.3.0334. Endocr Rev. 1998. PMID: 9626554 Review.
-
Prolactin receptor gene transcriptional control, regulatory modalities relevant to breast cancer resistance and invasiveness.Front Endocrinol (Lausanne). 2022 Sep 15;13:949396. doi: 10.3389/fendo.2022.949396. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 36187116 Free PMC article. Review.
Cited by
-
Molecular mechanisms of prolactin and its receptor.Endocr Rev. 2012 Aug;33(4):504-25. doi: 10.1210/er.2011-1040. Epub 2012 May 10. Endocr Rev. 2012. PMID: 22577091 Free PMC article. Review.
-
Paradigm-shifters: phosphorylated prolactin and short prolactin receptors.J Mammary Gland Biol Neoplasia. 2008 Mar;13(1):69-79. doi: 10.1007/s10911-008-9072-x. Epub 2008 Jan 25. J Mammary Gland Biol Neoplasia. 2008. PMID: 18219563 Review.
-
Structural modeling of cytokine-receptor-JAK2 signaling complexes using AlphaFold Multimer.bioRxiv [Preprint]. 2023 Jun 14:2023.06.14.544971. doi: 10.1101/2023.06.14.544971. bioRxiv. 2023. Update in: J Chem Inf Model. 2023 Sep 25;63(18):5874-5895. doi: 10.1021/acs.jcim.3c00926. PMID: 37398331 Free PMC article. Updated. Preprint.
-
Structural Modeling of Cytokine-Receptor-JAK2 Signaling Complexes Using AlphaFold Multimer.J Chem Inf Model. 2023 Sep 25;63(18):5874-5895. doi: 10.1021/acs.jcim.3c00926. Epub 2023 Sep 11. J Chem Inf Model. 2023. PMID: 37694948 Free PMC article.
-
Orchestration of signaling by structural disorder in class 1 cytokine receptors.Cell Commun Signal. 2020 Aug 24;18(1):132. doi: 10.1186/s12964-020-00626-6. Cell Commun Signal. 2020. PMID: 32831102 Free PMC article.
References
-
- Neville MC, McFadden TB, Forsyth I. Hormonal regulation of mammary differentiation and milk secretion. J. Mammary Gland Biol. Neoplasia. 2002;7:49–66. - PubMed
-
- Goffin V, Binart N, Touraine P, Kelly PA. Prolactin: the new biology of an old hormone. Ann. Rev. Physiol. 2002;64:47–67. - PubMed
-
- Hu Z, Zhuang L, Maria L, Dufau ML. Multiple and Tissue-specific Promoter Control of Gonadal and Non-gonadal Prolactin Receptor Gene Expression. J. Biol. Chem. 1996;271:10242–10246. - PubMed
-
- Nagano M, Kelly PA. Tissue distribution and regulation of rat prolactin receptor gene expression. Quantitative analysis by polymerase chain reaction. J. Biol. Chem. 1994;269:13337–13345. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
