

Sucrose transporter StSUT4 from potato affects flowering, tuberization, and shade avoidance response
- PMID: 18083796
- PMCID: PMC2245842
- DOI: 10.1104/pp.107.112334
Sucrose transporter StSUT4 from potato affects flowering, tuberization, and shade avoidance response
Abstract
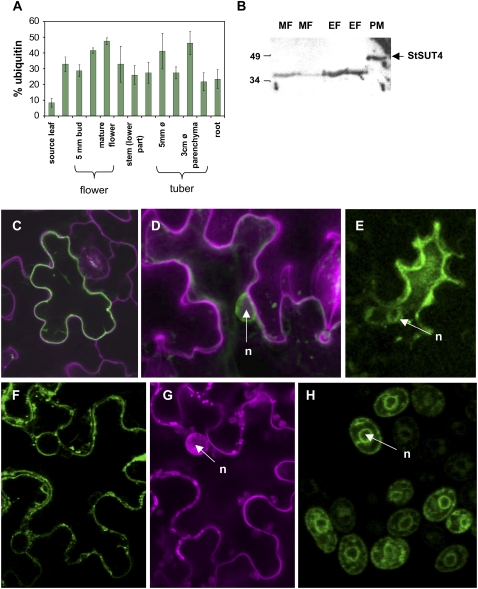
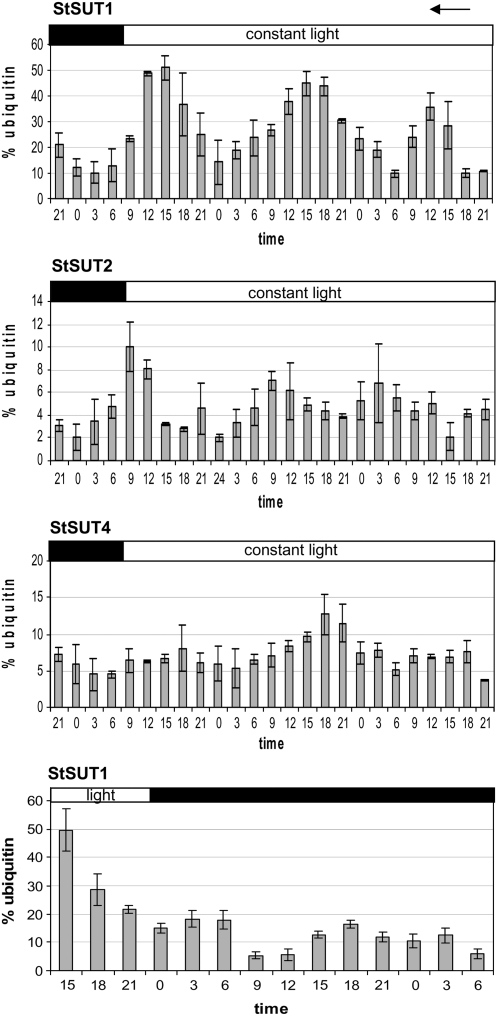
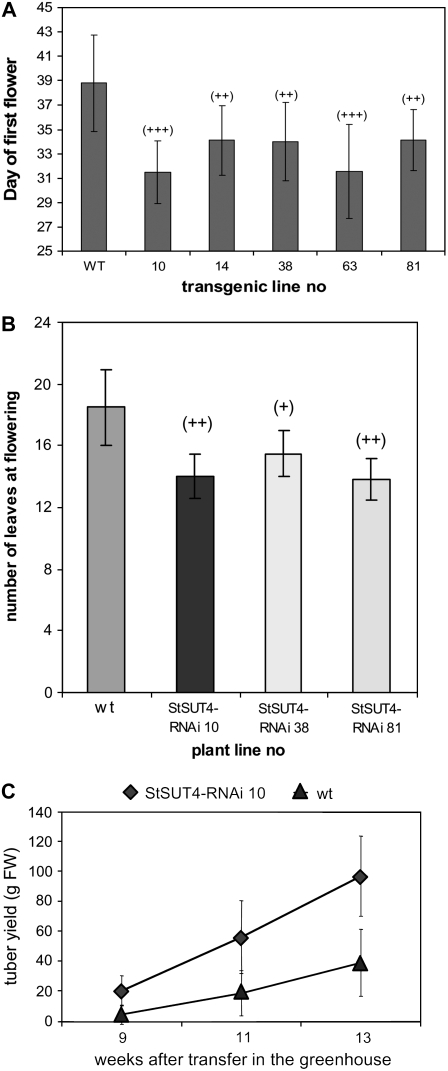
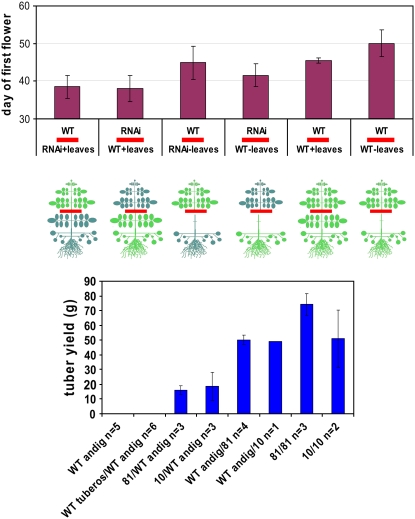
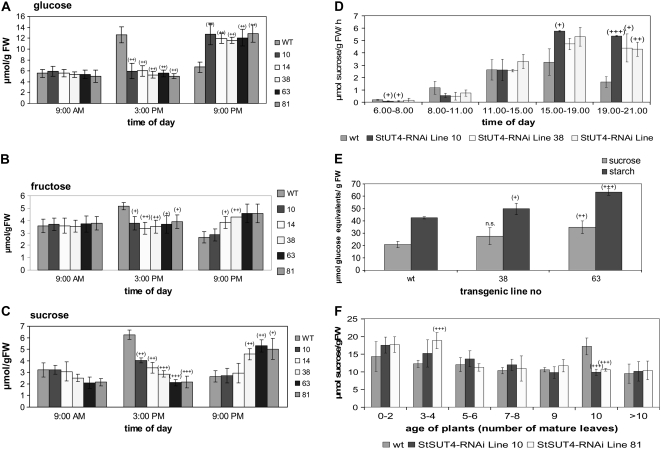
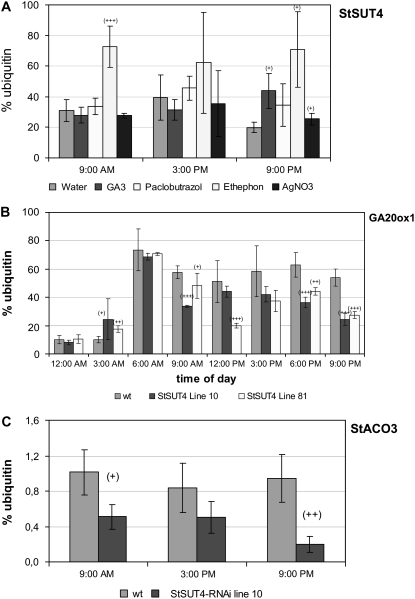
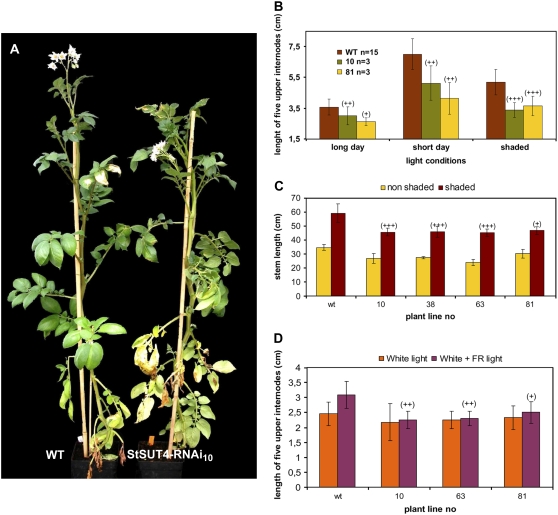
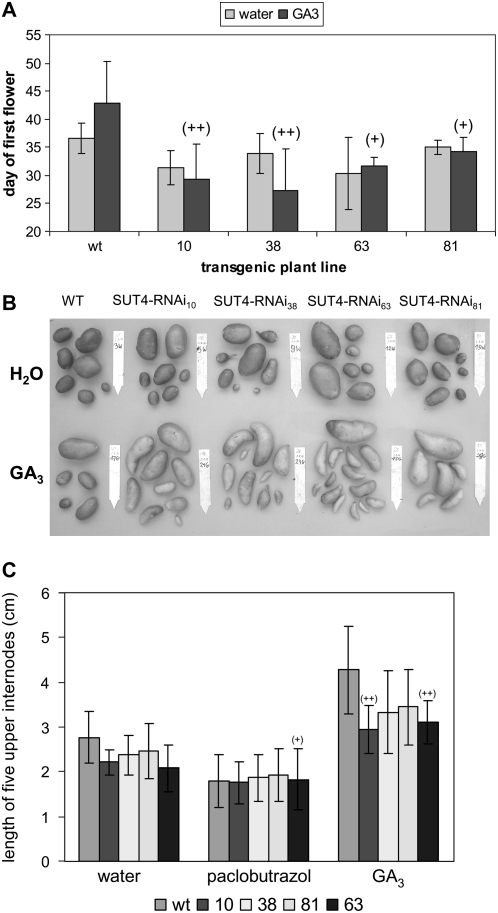
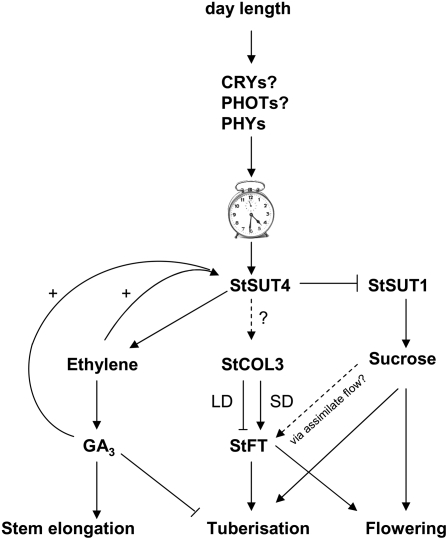
Sucrose (Suc) transporters belong to a large gene family. The physiological role of SUT1 proteins has been intensively investigated in higher plants, whereas that of SUT4 proteins is so far unknown. All three known Suc transporters from potato (Solanum tuberosum), SUT1, SUT2, and SUT4, are colocalized and their RNA levels not only follow a diurnal rhythm, but also oscillate in constant light. Here, we examined the physiological effects of transgenic potato plants on RNA interference (RNAi)-inactivated StSUT4 expression. The phenotype of StSUT4-RNAi plants includes early flowering, higher tuber production, and reduced sensitivity toward light enriched in far-red wavelength (i.e. in canopy shade). Inhibition of StSUT4 led to tuber production of the strict photoperiodic potato subsp. andigena even under noninductive long-day conditions. Accumulation of soluble sugars and Suc efflux from leaves of transgenic plants are modified in StSUT4-RNAi plants, leading to modified Suc levels in sink organs. StSUT4 expression of wild-type plants is induced by gibberellins and ethephon, and external supply of gibberellic acid leads to even more pronounced differences between wild-type and StSUT4-RNAi plants regarding tuber yield and internode elongation, indicating a reciprocal regulation of StSUT4 and gibberellins.
Figures

Similar articles
-
Expression Level of Mature miR172 in Wild Type and StSUT4-Silenced Plants of Solanum tuberosum Is Sucrose-Dependent.Int J Mol Sci. 2021 Feb 1;22(3):1455. doi: 10.3390/ijms22031455. Int J Mol Sci. 2021. PMID: 33535646 Free PMC article.
-
The sucrose transporter StSUT1 localizes to sieve elements in potato tuber phloem and influences tuber physiology and development.Plant Physiol. 2003 Jan;131(1):102-13. doi: 10.1104/pp.011676. Plant Physiol. 2003. PMID: 12529519 Free PMC article.
-
Photoperiodic regulation of the sucrose transporter StSUT4 affects the expression of circadian-regulated genes and ethylene production.Front Plant Sci. 2013 Feb 20;4:26. doi: 10.3389/fpls.2013.00026. eCollection 2013. Front Plant Sci. 2013. PMID: 23429841 Free PMC article.
-
Molecular signals that govern tuber development in potato.Int J Dev Biol. 2020;64(1-2-3):133-140. doi: 10.1387/ijdb.190132ab. Int J Dev Biol. 2020. PMID: 32659001 Review.
-
Key players associated with tuberization in potato: potential candidates for genetic engineering.Crit Rev Biotechnol. 2017 Nov;37(7):942-957. doi: 10.1080/07388551.2016.1274876. Epub 2017 Jan 18. Crit Rev Biotechnol. 2017. PMID: 28095718 Review.
Cited by
-
Over-expression of AtPAP2 in Camelina sativa leads to faster plant growth and higher seed yield.Biotechnol Biofuels. 2012 Apr 2;5:19. doi: 10.1186/1754-6834-5-19. Biotechnol Biofuels. 2012. PMID: 22472516 Free PMC article.
-
Drought response transcriptomes are altered in poplar with reduced tonoplast sucrose transporter expression.Sci Rep. 2016 Sep 19;6:33655. doi: 10.1038/srep33655. Sci Rep. 2016. PMID: 27641356 Free PMC article.
-
A critical appraisal of phloem-mobile signals involved in tuber induction.Front Plant Sci. 2013 Jul 16;4:253. doi: 10.3389/fpls.2013.00253. eCollection 2013. Front Plant Sci. 2013. PMID: 23882274 Free PMC article.
-
Metabolic engineering of raffinose-family oligosaccharides in the phloem reveals alterations in carbon partitioning and enhances resistance to green peach aphid.Front Plant Sci. 2013 Jul 19;4:263. doi: 10.3389/fpls.2013.00263. eCollection 2013. Front Plant Sci. 2013. PMID: 23882277 Free PMC article.
-
Molecular Evolution and Stress and Phytohormone Responsiveness of SUT Genes in Gossypium hirsutum.Front Genet. 2018 Oct 23;9:494. doi: 10.3389/fgene.2018.00494. eCollection 2018. Front Genet. 2018. PMID: 30405700 Free PMC article.
References
-
- Bernier G, Perilleux C (2005) A physiological overview of the genetics of flowering time control. Plant Biotechnol J 3 3–16 - PubMed
-
- Carrera E, Bou J, Garcia-Martinez JL, Prat S (2000) Changes in GA 20-oxidase gene expression strongly affect stem length, tuber induction and tuber yield of potato plants. Plant J 22 247–256 - PubMed
-
- Cerdan PD, Chory J (2003) Regulation of flowering time by light quality. Nature 423 881–885 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
