

doi: 10.1128/JB.01488-07.
Epub 2007 Dec 14.
Tn5 synaptic complex formation: role of transposase residue W450
Affiliations
- PMID: 18083803
- PMCID: PMC2238198
- DOI: 10.1128/JB.01488-07
Item in Clipboard
Tn5 synaptic complex formation: role of transposase residue W450
J Bacteriol.
2008 Feb.
Abstract
A series of Tn5 transposases (Tnp's) with mutations at the conserved amino acid position W450, which was structurally predicted to be important for synapsis, have been generated and characterized. This study demonstrates that W450 is involved in hydrophobic (and possibly aromatic) contacts within the Tnp monomer that negatively regulate synaptic complex formation.
Figures
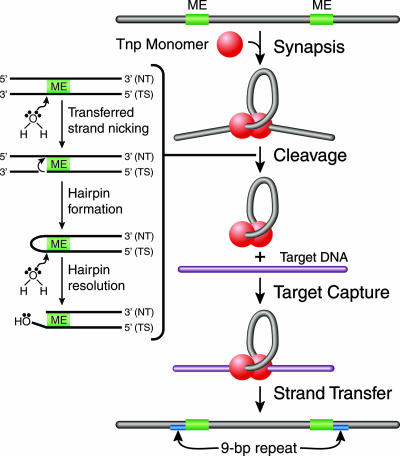
Tn5 transposition mechanism. Transposition is initiated by Tnp binding to the transposon-specific ESs and the formation of a highly ordered nucleoprotein complex (synaptic complex) through a process called synapsis. The synaptic complex contains two protomers of Tnp, which exist as a dimer, and two ESs. Catalytic cleavage occurs when an activated water molecule coordinated by Mg2+ nicks the transferred DNA strand (TS) on both sides of the transposon, through a nucleophilic attack, forming a 3′-hydroxyl group. The free 3′-hydroyxl group acts as a nucleophile and cleaves the nontransferred DNA strand (NT), forming a hairpin. A second activated water molecule resolves the hairpin, resulting in a double-stranded DNA cleavage product. The postcleavage synaptic complex is now free to bind to target DNA through target capture. The 3′-hydroxyl group of the transposon end attacks the phosphodiester backbone of target DNA during strand transfer. A 9-bp duplication in the target results, due to the staggered strand transfer reactions followed by DNA repair by host enzymes.
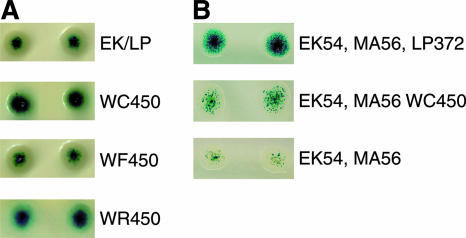
In vivo (papillation) transposition assay. Transposition levels were estimated by using the total number of blue papillae, and mutant Tnp's were compared to control Tnp to qualitatively assess transposase activity levels. (A) The W450F mutant Tnp (WF450) showed the same relative transposition activity as the control, indicating that W450 is involved in hydrophobic (and possibly aromatic) contacts. The W450C (WC450), W450S, and W450R (WR450) mutant Tnp's led to an ∼6-fold increase in transposition activity and proceeded to catalyze transposition (form blue papillae) ∼8 h before the control (EK/LP) (data not shown). (B) Transposition activity levels of the W450C mutant Tnp (lacking L372P) and the control were compared. The W450C mutant Tnp was shown to be less active in transposition than control Tnp, but the mutation at position 450 still led to a general increase in transposition activity compared to that of Tnp containing the E54K (EK54) and M56A (MA56) mutations. This finding indicates that the increase in transposition activity conferred by mutations at position 450 is not dependent on the presence of the L372P (LP372) mutation.
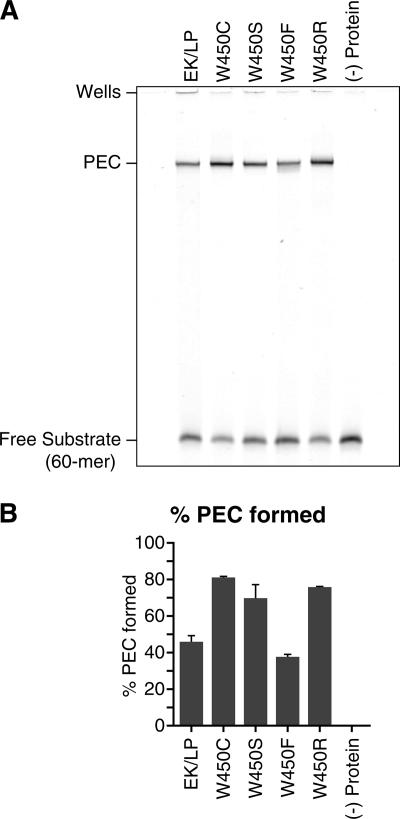
PEC formation assay. (A) PECs were formed by incubating Tnp with double-stranded, fluorescently labeled DNA oligomers (60-mers) for 1 h at 37°C as previously described, with the addition of 100 μg of tRNA/ml to the reaction buffer (13). Products were visualized via 6% native PAGE. These experiments were repeated three times, and the gel shown is representative of all of these experiments. EK/LP, control Tnp; (−) protein, no protein. (B) PEC formation was quantified by comparing the amount of labeled DNA in the PEC to the total amount of labeled DNA. These experiments were repeated three times, and the results shown are representative of all of these experiments. PEC formation was enhanced for W450C, W450S, and W450R mutant Tnp's, which formed 81, 70, and 76% PECs, respectively, in contrast to the control Tnp, which formed 46% PECs. These differences represent increases of 1.8-, 1.5-, and 1.8-fold, respectively. The W450F mutant Tnp formed 38% PECs, which represents an 18% reduction compared to the control Tnp. Increases in PEC formation were most likely due to the removal of the hydrophobic (and possibly aromatic) contacts between amino acid W450 and its binding partners, presumably on the N-terminal portion of the Tnp monomer. In this model, the removal of these contacts shifts the conformation equilibrium of the Tnp monomer away from the inhibitory conformation.
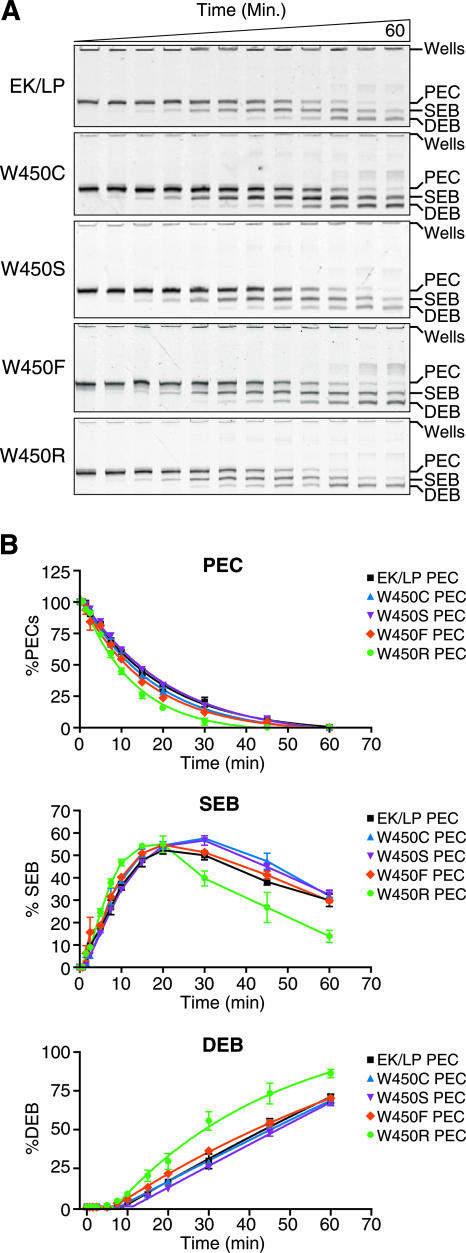
Cleavage assay. (A) Cleavage activity was assessed by forming PECs with fluorescently labeled DNA oligomers (60-mer) at 37°C for 1 h as previously described, with the addition of 100 μg of tRNA/ml to the reaction buffer (13). After complex formation, Mg2+ was added to a final concentration of 10 mM to catalyze the cleavage mechanism. Cleavage results in two products: the SEB and the DEB. Cleavage products, along with unreacted PEC, were visualized via 6% native PAGE. The time course proceeded for 1 h. These experiments were repeated three times, and the gel shown is representative of all of these experiments. EK/LP, control Tnp. (B) The percentage of labeled oligonucleotide in each complex (the PEC, SEB, or DEB) relative to the total amount of the substrate contained in the complex was determined. With the exception of the W450R mutant Tnp, the Tnp's with mutations at position W450 behaved similarly to the control Tnp. The mutant Tnp forms had approximately the same rates and total yields of PEC cleavage, SEB formation and destruction, and DEB formation. The W450R mutant Tnp led to an increase in the rate of cleavage over that of the control, forming SEB complexes and converting SEB complexes into DEB complexes faster than the control. This increased rate of cleavage was repeatable over all three separate experiments. The data indicate that position W450 does not play a large role in the cleavage mechanism but that the substitution of a large charged amino acid leads to hyperactivity, possibly through steric interference with the W450 binding partners.
Similar articles
-
Site-directed mutagenesis studies of tn5 transposase residues involved in synaptic complex formation.J Bacteriol. 2007 Oct;189(20):7436-41. doi: 10.1128/JB.00524-07. Epub 2007 Aug 10. J Bacteriol. 2007. PMID: 17693501 Free PMC article.
-
Tn5 transposase loops DNA in the absence of Tn5 transposon end sequences.Mol Microbiol. 2006 Dec;62(6):1558-68. doi: 10.1111/j.1365-2958.2006.05471.x. Mol Microbiol. 2006. PMID: 17074070
-
Mutation of Tn5 transposase beta-loop residues affects all steps of Tn5 transposition: the role of conformational changes in Tn5 transposition.Biochemistry. 2006 Dec 26;45(51):15552-62. doi: 10.1021/bi061227v. Epub 2006 Dec 5. Biochemistry. 2006. PMID: 17176076 Free PMC article.
-
Tn5 as a model for understanding DNA transposition.Mol Microbiol. 2003 Mar;47(5):1199-206. doi: 10.1046/j.1365-2958.2003.03382.x. Mol Microbiol. 2003. PMID: 12603728 Review.
-
Structure/function insights into Tn5 transposition.Curr Opin Struct Biol. 2004 Feb;14(1):50-7. doi: 10.1016/j.sbi.2004.01.008. Curr Opin Struct Biol. 2004. PMID: 15102449 Review.
References
-
- Betermier, M., R. Alazard, F. Ragueh, E. Roulet, A. Toussaint, and M. Chandler. 1987. Phage Mu transposase: deletion of the carboxy-terminal end does not abolish DNA-binding activity. Mol. Gen. Genet. 21077-85. - PubMed
-
- Bhasin, A., I. Y. Goryshin, M. Steiniger-White, D. York, and W. S. Reznikoff. 2000. Characterization of a Tn5 pre-cleavage synaptic complex. J. Mol. Biol. 30249-63. - PubMed
-
- Davies, D. R., I. Y. Goryshin, W. S. Reznikoff, and I. Rayment. 2000. Three-dimensional structure of the Tn5 synaptic complex transposition intermediate. Science 28977-85. - PubMed
-
- Davies, D. R., L. Mahnke Braam, W. S. Reznikoff, and I. Rayment. 1999. The three-dimensional structure of a Tn5 transposase-related protein determined to 2.9-Å resolution. J. Biol. Chem. 27411904-11913. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
