

Interaction between two putative glycosyltransferases is required for glycosylation of a serine-rich streptococcal adhesin
- PMID: 18083807
- PMCID: PMC2238222
- DOI: 10.1128/JB.01078-07
Interaction between two putative glycosyltransferases is required for glycosylation of a serine-rich streptococcal adhesin
Abstract
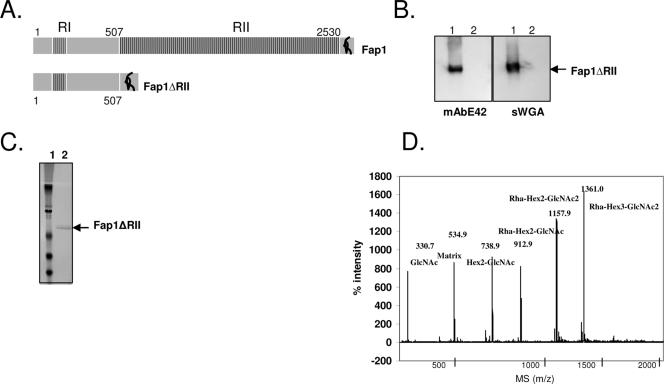
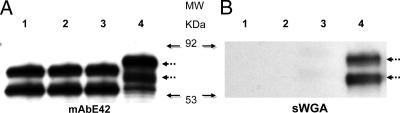
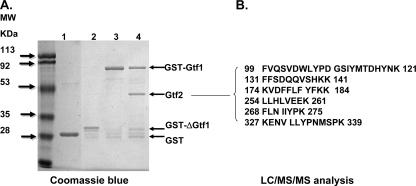
Fap1, a serine-rich glycoprotein, is essential for fimbrial biogenesis and biofilm formation of Streptococcus parasanguinis (formerly S. parasanguis). Fap1-like proteins are conserved in many streptococci and staphylococci and have been implicated in bacterial virulence. Fap1 contains two serine-rich repeat regions that are modified by O-linked glycosylation. A seven-gene cluster has been identified, and this cluster is implicated in Fap1 biogenesis. In this study, we investigated the initial step of Fap1 glycosylation by using a recombinant Fap1 as a model. This recombinant molecule has the same monosaccharide composition profile as the native Fap1 protein. Glycosyl linkage analyses indicated that N-acetylglucosamine (GlcNAc) is among the first group of sugar residues transferred to the Fap1 peptide. Two putative glycosyltransferases, Gtf1 and Gtf2, were essential for the glycosylation of Fap1 with GlcNAc-containing oligosaccharide(s) in both S. parasanguinis as well as in the Fap1 glycosylation system in Escherichia coli. Yeast two-hybrid analysis as well as in vitro and in vivo glutathione S-transferase pull-down assays demonstrated the two putative glycosyltransferases interacted with each other. The interaction domain was mapped to an N-terminal region of Gtf1 that was required for the Fap1 glycosylation. The data in this study suggested that the formation of the Gtf1 and Gtf2 complex was required for the initiation of the Fap1 glycosylation and that the N-terminal region of Gtf1 was necessary.
Figures



Similar articles
-
A molecular chaperone mediates a two-protein enzyme complex and glycosylation of serine-rich streptococcal adhesins.J Biol Chem. 2011 Oct 7;286(40):34923-31. doi: 10.1074/jbc.M111.239350. Epub 2011 Aug 23. J Biol Chem. 2011. PMID: 21862581 Free PMC article.
-
A novel glucosyltransferase is required for glycosylation of a serine-rich adhesin and biofilm formation by Streptococcus parasanguinis.J Biol Chem. 2010 Apr 16;285(16):12140-8. doi: 10.1074/jbc.M109.066928. Epub 2010 Feb 17. J Biol Chem. 2010. PMID: 20164186 Free PMC article.
-
A conserved domain of previously unknown function in Gap1 mediates protein-protein interaction and is required for biogenesis of a serine-rich streptococcal adhesin.Mol Microbiol. 2008 Dec;70(5):1094-104. doi: 10.1111/j.1365-2958.2008.06456.x. Epub 2008 Sep 30. Mol Microbiol. 2008. PMID: 18826412 Free PMC article.
-
Glycosylation and biogenesis of a family of serine-rich bacterial adhesins.Microbiology (Reading). 2009 Feb;155(Pt 2):317-327. doi: 10.1099/mic.0.025221-0. Microbiology (Reading). 2009. PMID: 19202081 Review.
-
Glycosyltransferase-mediated Sweet Modification in Oral Streptococci.J Dent Res. 2015 May;94(5):659-65. doi: 10.1177/0022034515574865. Epub 2015 Mar 9. J Dent Res. 2015. PMID: 25755271 Free PMC article. Review.
Cited by
-
Serine-rich repeat proteins: well-known yet little-understood bacterial adhesins.J Bacteriol. 2024 Jan 25;206(1):e0024123. doi: 10.1128/jb.00241-23. Epub 2023 Nov 17. J Bacteriol. 2024. PMID: 37975670 Free PMC article. Review.
-
A molecular chaperone mediates a two-protein enzyme complex and glycosylation of serine-rich streptococcal adhesins.J Biol Chem. 2011 Oct 7;286(40):34923-31. doi: 10.1074/jbc.M111.239350. Epub 2011 Aug 23. J Biol Chem. 2011. PMID: 21862581 Free PMC article.
-
Modification of Streptococcus mutans Cnm by PgfS contributes to adhesion, endothelial cell invasion, and virulence.J Bacteriol. 2014 Aug;196(15):2789-97. doi: 10.1128/JB.01783-14. Epub 2014 May 16. J Bacteriol. 2014. PMID: 24837294 Free PMC article.
-
Transcriptional organization of pneumococcal psrP-secY2A2 and impact of GtfA and GtfB deletion on PsrP-associated virulence properties.Microbes Infect. 2017 Jun;19(6):323-333. doi: 10.1016/j.micinf.2017.04.001. Epub 2017 Apr 10. Microbes Infect. 2017. PMID: 28408270 Free PMC article.
-
Interactions in bacterial biofilm development: a structural perspective.Curr Protein Pept Sci. 2012 Dec;13(8):739-55. doi: 10.2174/138920312804871166. Curr Protein Pept Sci. 2012. PMID: 23305361 Free PMC article. Review.
References
-
- Carlsson, J., H. Grahnen, G. Jonsson, and S. Wikner. 1970. Establishment of Streptococcus sanguis in the mouths of infants. Arch. Oral Biol. 151143-1148. - PubMed
-
- Chen, Q., H. Wu, and P. M. Fives-Taylor. 2004. Investigating the role of secA2 in secretion and glycosylation of a fimbrial adhesin in Streptococcus parasanguis FW213. Mol. Microbiol. 53843-856. - PubMed
-
- Chen, Q., H. Wu, R. Kumar, Z. Peng, and P. M. Fives-Taylor. 2006. SecA2 is distinct from SecA in immunogenic specificity, subcellular distribution and requirement for membrane anchoring in Streptococcus parasanguis. FEMS Microbiol. Lett. 264174-181. - PubMed
-
- Cole, R. M., G. B. Calandra, E. Huff, and K. M. Nugent. 1976. Attributes of potential utility in differentiating among “group H” streptococci or Streptococcus sanguis. J. Dent. Res. 55A142-A153. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
