

Clostridium perfringens spore germination: characterization of germinants and their receptors
- PMID: 18083820
- PMCID: PMC2238220
- DOI: 10.1128/JB.01748-07
Clostridium perfringens spore germination: characterization of germinants and their receptors
Abstract
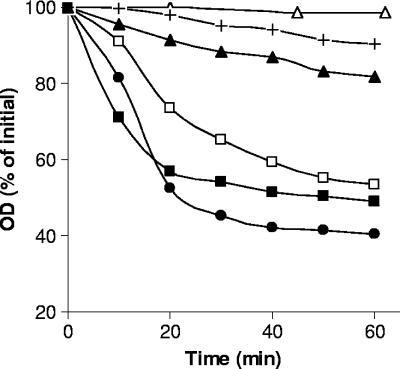
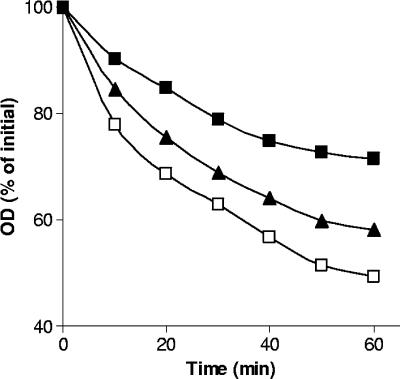
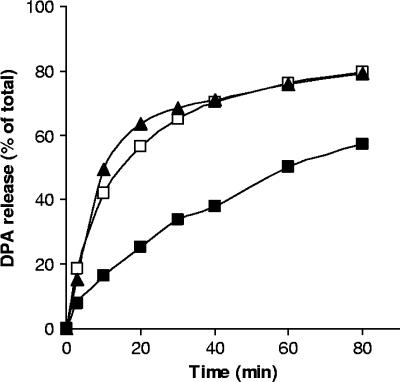
Clostridium perfringens food poisoning is caused by type A isolates carrying a chromosomal enterotoxin (cpe) gene (C-cpe), while C. perfringens-associated non-food-borne gastrointestinal (GI) diseases are caused by isolates carrying a plasmid-borne cpe gene (P-cpe). C. perfringens spores are thought to be the important infectious cell morphotype, and after inoculation into a suitable host, these spores must germinate and return to active growth to cause GI disease. We have found differences in the germination of spores of C-cpe and P-cpe isolates in that (i) while a mixture of L-asparagine and KCl was a good germinant for spores of C-cpe and P-cpe isolates, KCl and, to a lesser extent, L-asparagine triggered spore germination in C-cpe isolates only; and (ii) L-alanine or L-valine induced significant germination of spores of P-cpe but not C-cpe isolates. Spores of a gerK mutant of a C-cpe isolate in which two of the proteins of a spore nutrient germinant receptor were absent germinated slower than wild-type spores with KCl, did not germinate with L-asparagine, and germinated poorly compared to wild-type spores with the nonnutrient germinants dodecylamine and a 1:1 chelate of Ca2+ and dipicolinic acid. In contrast, spores of a gerAA mutant of a C-cpe isolate that lacked another component of a nutrient germinant receptor germinated at the same rate as that of wild-type spores with high concentrations of KCl, although they germinated slightly slower with a lower KCl concentration, suggesting an auxiliary role for GerAA in C. perfringens spore germination. In sum, this study identified nutrient germinants for spores of both C-cpe and P-cpe isolates of C. perfringens and provided evidence that proteins encoded by the gerK operon are required for both nutrient-induced and non-nutrient-induced spore germination.
Figures








Similar articles
-
l-lysine (pH 6.0) induces germination of spores of Clostridium perfringens type F isolates carrying chromosomal or plasmid-borne enterotoxin gene.Microb Pathog. 2018 Oct;123:227-232. doi: 10.1016/j.micpath.2018.07.022. Epub 2018 Jul 18. Microb Pathog. 2018. PMID: 30031038
-
Bicarbonate and amino acids are co-germinants for spores of Clostridium perfringens type A isolates carrying plasmid-borne enterotoxin gene.Food Microbiol. 2018 Feb;69:64-71. doi: 10.1016/j.fm.2017.06.020. Epub 2017 Jul 26. Food Microbiol. 2018. PMID: 28941910
-
Characterization of germinants and their receptors for spores of non-food-borne Clostridium perfringens strain F4969.Microbiology (Reading). 2016 Nov;162(11):1972-1983. doi: 10.1099/mic.0.000378. Epub 2016 Sep 29. Microbiology (Reading). 2016. PMID: 27692042
-
The biology and pathogenicity of Clostridium perfringens type F: a common human enteropathogen with a new(ish) name.Microbiol Mol Biol Rev. 2024 Sep 26;88(3):e0014023. doi: 10.1128/mmbr.00140-23. Epub 2024 Jun 12. Microbiol Mol Biol Rev. 2024. PMID: 38864615 Free PMC article. Review.
-
Enterotoxigenic Clostridium perfringens: detection and identification.Microbes Environ. 2012;27(4):343-9. doi: 10.1264/jsme2.me12002. Epub 2012 Apr 14. Microbes Environ. 2012. PMID: 22504431 Free PMC article. Review.
Cited by
-
Cwl0971, a novel peptidoglycan hydrolase, plays pleiotropic roles in Clostridioides difficile R20291.Environ Microbiol. 2021 Sep;23(9):5222-5238. doi: 10.1111/1462-2920.15529. Epub 2021 Apr 24. Environ Microbiol. 2021. PMID: 33893759 Free PMC article.
-
The small acid-soluble proteins of Clostridioides difficile regulate sporulation in a SpoIVB2-dependent manner.PLoS Pathog. 2024 Aug 30;20(8):e1012507. doi: 10.1371/journal.ppat.1012507. eCollection 2024 Aug. PLoS Pathog. 2024. PMID: 39213448 Free PMC article.
-
Clostridioides difficile Biology: Sporulation, Germination, and Corresponding Therapies for C. difficile Infection.Front Cell Infect Microbiol. 2018 Feb 8;8:29. doi: 10.3389/fcimb.2018.00029. eCollection 2018. Front Cell Infect Microbiol. 2018. PMID: 29473021 Free PMC article. Review.
-
Spore Cortex Hydrolysis Precedes Dipicolinic Acid Release during Clostridium difficile Spore Germination.J Bacteriol. 2015 Jul;197(14):2276-83. doi: 10.1128/JB.02575-14. Epub 2015 Apr 27. J Bacteriol. 2015. PMID: 25917906 Free PMC article.
-
The role of supplemental glycine in establishing a subclinical necrotic enteritis challenge model in broiler chickens.Anim Nutr. 2017 Sep;3(3):266-270. doi: 10.1016/j.aninu.2017.05.004. Epub 2017 May 25. Anim Nutr. 2017. PMID: 29767149 Free PMC article.
References
-
- Alberto, F., V. Broussolle, D. R. Mason, F. Carlin, and M. W. Peck. 2003. Variability in spore germination response by strains of proteolytic Clostridium botulinum types A, B and F. Lett. Appl. Microbiol. 3641-45. - PubMed
-
- Bannam, T. L., and J. I. Rood. 1993. Clostridium perfringens-Escherichia coli shuttle vectors that carry single antibiotic resistance determinants. Plasmid 29233-235. - PubMed
-
- Broussolle, V., F. Alberto, C. A. Shearman, D. R. Mason, L. Botella, C. Nguyen-The, M. W. Peck, and F. Carlin. 2002. Molecular and physiological characterization of spore germination in Clostridium botulinum and C. sporogenes. Anaerobe 889-100.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
