

The E. coli RhlE RNA helicase regulates the function of related RNA helicases during ribosome assembly
- PMID: 18083833
- PMCID: PMC2212244
- DOI: 10.1261/rna.800308
The E. coli RhlE RNA helicase regulates the function of related RNA helicases during ribosome assembly
Abstract
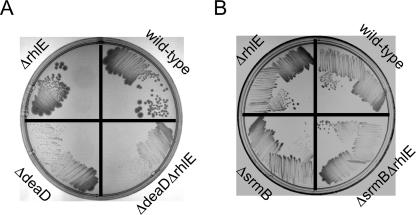
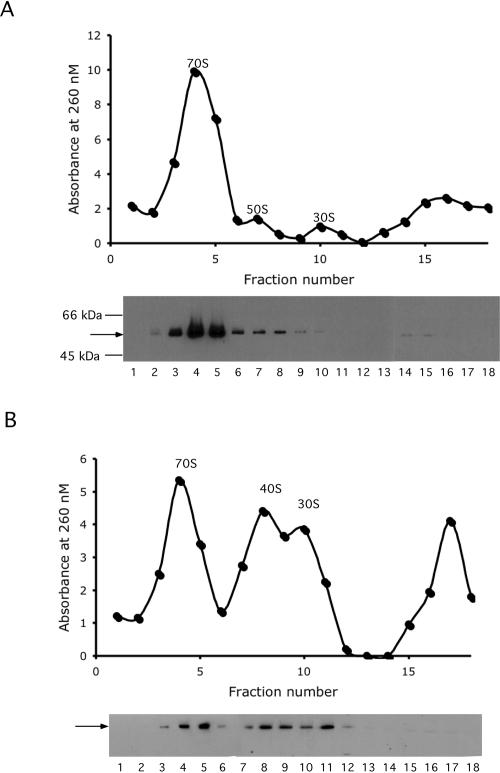
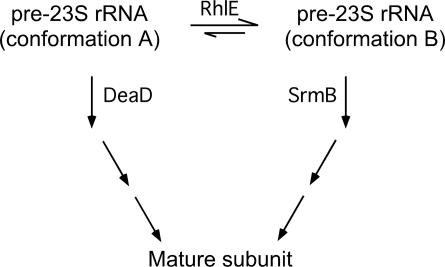
Escherichia coli contains five members of the DEAD-box RNA helicase family, a ubiquitous class of proteins characterized by their ability to unwind RNA duplexes. Although four of these proteins have been implicated in RNA turnover or ribosome biogenesis, no cellular function for the RhlE DEAD-box protein has been described as yet. During an analysis of the cold-sensitive growth defect of a strain lacking the DeaD/CsdA RNA helicase, rhlE plasmids were identified from a chromosomal library as multicopy suppressors of the growth defect. Remarkably, when tested for allele specificity, RhlE overproduction was found to exacerbate the cold-sensitive growth defect of a strain that lacks the SrmB RNA helicase. Moreover, the absence of RhlE exacerbated or alleviated the cold-sensitive defect of deaD or srmB strains, respectively. Primer extension and ribosome analysis indicated that RhlE regulates the accumulation of immature ribosomal RNA or ribosome precursors when deaD or srmB strains are grown at low temperatures. By using an epitope-tagged version of RhlE, the majority of RhlE in cell extracts was found to cosediment with ribosome-containing fractions. Since both DeaD and SrmB have been recently shown to function in ribosome assembly, these findings suggests that rhlE genetically interacts with srmB and deaD to modulate their function during ribosome maturation. On the basis of the available evidence, I propose that RhlE is a novel ribosome assembly factor, which plays a role in the interconversion of ribosomal RNA-folding intermediates that are further processed by DeaD or SrmB during ribosome maturation.
Figures



Similar articles
-
Identification of the sites of action of SrmB, a DEAD-box RNA helicase involved in Escherichia coli ribosome assembly.Mol Microbiol. 2011 Oct;82(2):300-11. doi: 10.1111/j.1365-2958.2011.07779.x. Epub 2011 Aug 22. Mol Microbiol. 2011. PMID: 21859437
-
Association of the Cold Shock DEAD-Box RNA Helicase RhlE to the RNA Degradosome in Caulobacter crescentus.J Bacteriol. 2017 Jun 13;199(13):e00135-17. doi: 10.1128/JB.00135-17. Print 2017 Jul 1. J Bacteriol. 2017. PMID: 28396352 Free PMC article.
-
SrmB, a DEAD-box helicase involved in Escherichia coli ribosome assembly, is specifically targeted to 23S rRNA in vivo.Nucleic Acids Res. 2009 Oct;37(19):6540-9. doi: 10.1093/nar/gkp685. Epub 2009 Sep 4. Nucleic Acids Res. 2009. PMID: 19734346 Free PMC article.
-
Assembly of bacterial ribosomes.Annu Rev Biochem. 2011;80:501-26. doi: 10.1146/annurev-biochem-062608-160432. Annu Rev Biochem. 2011. PMID: 21529161 Review.
-
Functions of DEAD-box proteins in bacteria: current knowledge and pending questions.Biochim Biophys Acta. 2013 Aug;1829(8):866-77. doi: 10.1016/j.bbagrm.2013.01.012. Epub 2013 Feb 13. Biochim Biophys Acta. 2013. PMID: 23415794 Review.
Cited by
-
Antibacterial Activity of Rainbow Trout Plasma: In Vitro Assays and Proteomic Analysis.Animals (Basel). 2023 Nov 18;13(22):3565. doi: 10.3390/ani13223565. Animals (Basel). 2023. PMID: 38003182 Free PMC article.
-
Time course of large ribosomal subunit assembly in E. coli cells overexpressing a helicase inactive DbpA protein.RNA. 2016 Jul;22(7):1055-64. doi: 10.1261/rna.055137.115. Epub 2016 May 18. RNA. 2016. PMID: 27194011 Free PMC article.
-
Role of ribosome assembly in Escherichia coli ribosomal RNA degradation.Nucleic Acids Res. 2018 Nov 16;46(20):11048-11060. doi: 10.1093/nar/gky808. Nucleic Acids Res. 2018. PMID: 30219894 Free PMC article.
-
A DEAD-box protein regulates ribosome assembly through control of ribosomal protein synthesis.Nucleic Acids Res. 2019 Sep 5;47(15):8193-8206. doi: 10.1093/nar/gkz502. Nucleic Acids Res. 2019. PMID: 31188443 Free PMC article.
-
Deconstructing ribosome construction.Trends Biochem Sci. 2009 May;34(5):256-63. doi: 10.1016/j.tibs.2009.01.011. Epub 2009 Apr 17. Trends Biochem Sci. 2009. PMID: 19376708 Free PMC article. Review.
References
-
- Aiba, H., Adhya, S., de Crombrugghe, B. Evidence for two functional gal promoters in intact Escherichia coli cells. J. Biol. Chem. 1981;256:11905–11910. - PubMed
-
- Bizebard, T., Ferlenghi, I., Iost, I., Dreyfus, M. Studies on three E. coli DEAD-box helicases point to an unwinding mechanism different from that of model DNA helicases. Biochemistry. 2004;43:7857–7866. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous