

Modulation of utrophin A mRNA stability in fast versus slow muscles via an AU-rich element and calcineurin signaling
- PMID: 18084024
- PMCID: PMC2241908
- DOI: 10.1093/nar/gkm1107
Modulation of utrophin A mRNA stability in fast versus slow muscles via an AU-rich element and calcineurin signaling
Abstract
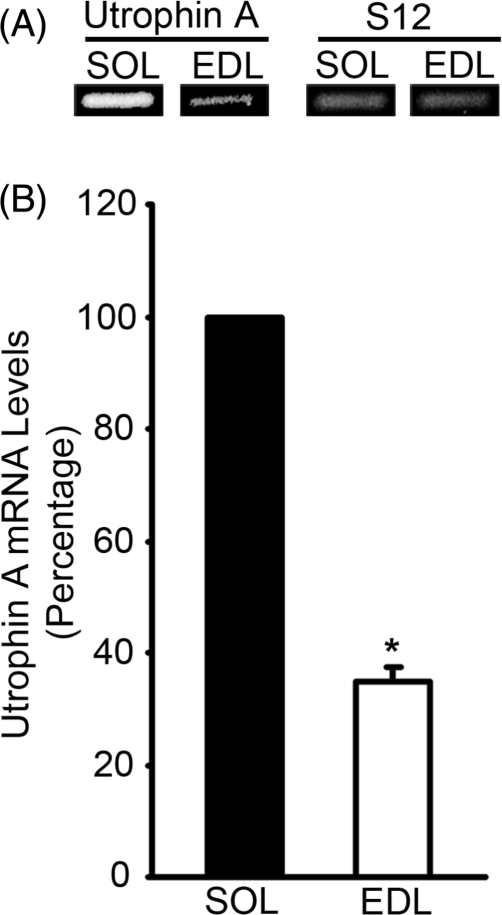
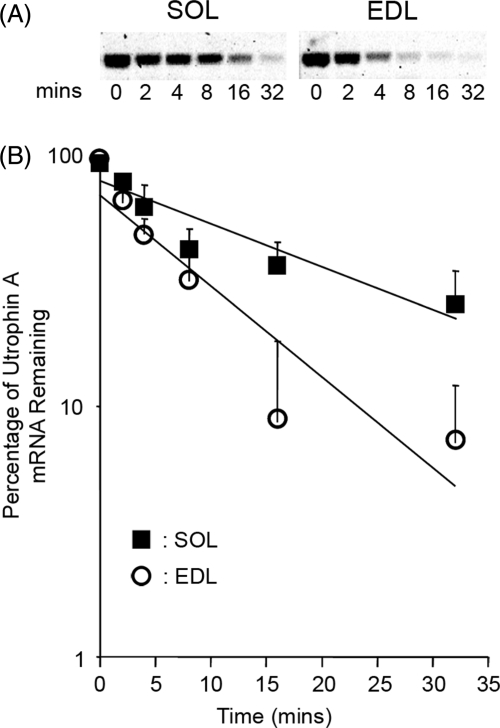
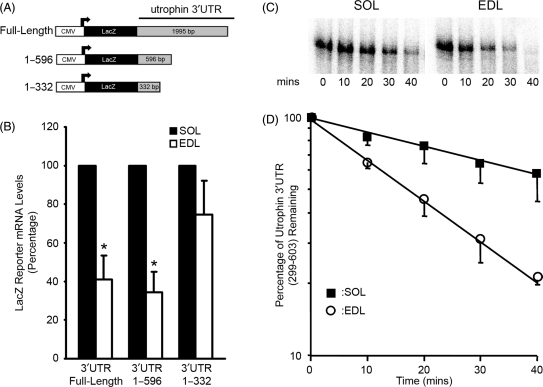
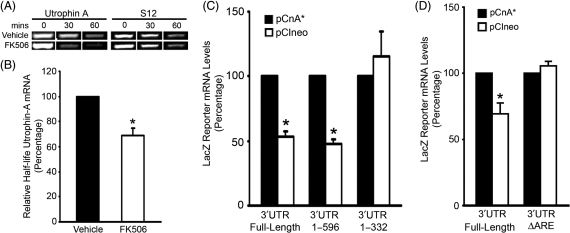
We examined the role of post-transcriptional mechanisms in controlling utrophin A mRNA expression in slow versus fast skeletal muscles. First, we determined that the half-life of utrophin A mRNA is significantly shorter in the presence of proteins isolated from fast muscles. Direct plasmid injection experiments using reporter constructs containing the full-length or truncated variants of the utrophin 3'UTR into slow soleus and fast extensor digitorum longus muscles revealed that a region of 265 nucleotides is sufficient to confer lower levels of reporter mRNA in fast muscles. Further analysis of this region uncovered a conserved AU-rich element (ARE) that suppresses expression of reporter mRNAs in cultured muscle cells. Moreover, stability of reporter mRNAs fused to the utrophin full-length 3'UTR was lower in the presence of fast muscle protein extracts. This destabilization effect seen in vivo was lost upon deletion of the conserved ARE. Finally, we observed that calcineurin signaling affects utrophin A mRNA stability through the conserved ARE. These results indicate that ARE-mediated mRNA decay is a key mechanism that regulates expression of utrophin A mRNA in slow muscle fibers. This is the first demonstration of ARE-mediated mRNA decay regulating the expression of a gene associated with the slow myogenic program.
Figures



Similar articles
-
Increased expression of utrophin in a slow vs. a fast muscle involves posttranscriptional events.Am J Physiol Cell Physiol. 2001 Oct;281(4):C1300-9. doi: 10.1152/ajpcell.2001.281.4.C1300. Am J Physiol Cell Physiol. 2001. PMID: 11546668
-
Activation of p38 signaling increases utrophin A expression in skeletal muscle via the RNA-binding protein KSRP and inhibition of AU-rich element-mediated mRNA decay: implications for novel DMD therapeutics.Hum Mol Genet. 2013 Aug 1;22(15):3093-111. doi: 10.1093/hmg/ddt165. Epub 2013 Apr 10. Hum Mol Genet. 2013. PMID: 23575223
-
Expression of utrophin A mRNA correlates with the oxidative capacity of skeletal muscle fiber types and is regulated by calcineurin/NFAT signaling.Proc Natl Acad Sci U S A. 2003 Jun 24;100(13):7791-6. doi: 10.1073/pnas.0932671100. Epub 2003 Jun 13. Proc Natl Acad Sci U S A. 2003. PMID: 12808150 Free PMC article.
-
AU-rich elements and associated factors: are there unifying principles?Nucleic Acids Res. 2006 Jan 3;33(22):7138-50. doi: 10.1093/nar/gki1012. Print 2005. Nucleic Acids Res. 2006. PMID: 16391004 Free PMC article. Review.
-
Multiple regulatory events controlling the expression and localization of utrophin in skeletal muscle fibers: insights into a therapeutic strategy for Duchenne muscular dystrophy.J Physiol Paris. 2002 Jan-Mar;96(1-2):31-42. doi: 10.1016/s0928-4257(01)00078-x. J Physiol Paris. 2002. PMID: 11755781 Review.
Cited by
-
High-throughput identification of post-transcriptional utrophin up-regulators for Duchenne muscle dystrophy (DMD) therapy.Sci Rep. 2020 Feb 7;10(1):2132. doi: 10.1038/s41598-020-58737-6. Sci Rep. 2020. PMID: 32034254 Free PMC article.
-
A global profiling of uncapped mRNAs under cold stress reveals specific decay patterns and endonucleolytic cleavages in Brachypodium distachyon.Genome Biol. 2013 Aug 30;14(8):R92. doi: 10.1186/gb-2013-14-8-r92. Genome Biol. 2013. PMID: 24000894 Free PMC article.
-
Second-generation compound for the modulation of utrophin in the therapy of DMD.Hum Mol Genet. 2015 Aug 1;24(15):4212-24. doi: 10.1093/hmg/ddv154. Epub 2015 May 1. Hum Mol Genet. 2015. PMID: 25935002 Free PMC article.
-
The era of genomic medicine.Clin Med (Lond). 2013 Dec;13(6):594-601. doi: 10.7861/clinmedicine.13-6-594. Clin Med (Lond). 2013. PMID: 24298109 Free PMC article. Review.
-
Alternative utrophin mRNAs contribute to phenotypic differences between dystrophin-deficient mice and Duchenne muscular dystrophy.FEBS Lett. 2018 Jun;592(11):1856-1869. doi: 10.1002/1873-3468.13099. Epub 2018 May 30. FEBS Lett. 2018. PMID: 29772070 Free PMC article.
References
-
- Krag TO, Gyrd-Hansen M, Khurana TS. Harnessing the potential of dystrophin-related proteins for ameliorating Duchenne's muscular dystrophy. Acta Physiol. Scand. 2001;171:349–358. - PubMed
-
- Blake DJ, Weir A, Newey SE, Davies KE. Function and genetics of dystrophin and dystrophin-related proteins in muscle. Physiol. Rev. 2002;82:291–329. - PubMed
-
- Khurana TS, Hoffman EP, Kunkel LM. Identification of a chromosome 6-encoded dystrophin-related protein. J. Biol. Chem. 1990;265:16717–16720. - PubMed
-
- Nguyen TM, Ellis JM, Love DR, Davies KE, Gatter KC, Dickson G, Morris GE. Localization of the DMDL gene-encoded dystrophin-related protein using a panel of nineteen monoclonal antibodies: presence at neuromuscular junctions, in the sarcolemma of dystrophic skeletal muscle, in vascular and other smooth muscles, and in proliferating brain cell lines. J. Cell Biol. 1991;115:1695–1700. - PMC - PubMed
-
- Ohlendieck K, Ervasti JM, Matsumura K, Kahl SD, Leveille CJ, Campbell KP. Dystrophin-related protein is localized to neuromuscular junctions of adult skeletal muscle. Neuron. 1991;7:499–508. - PubMed
