

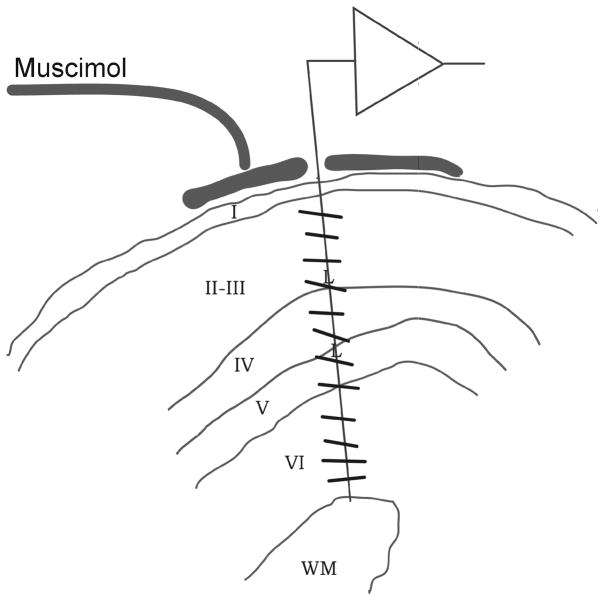
On and off domains of geniculate afferents in cat primary visual cortex
- PMID: 18084287
- PMCID: PMC2556869
- DOI: 10.1038/nn2029
On and off domains of geniculate afferents in cat primary visual cortex
Abstract
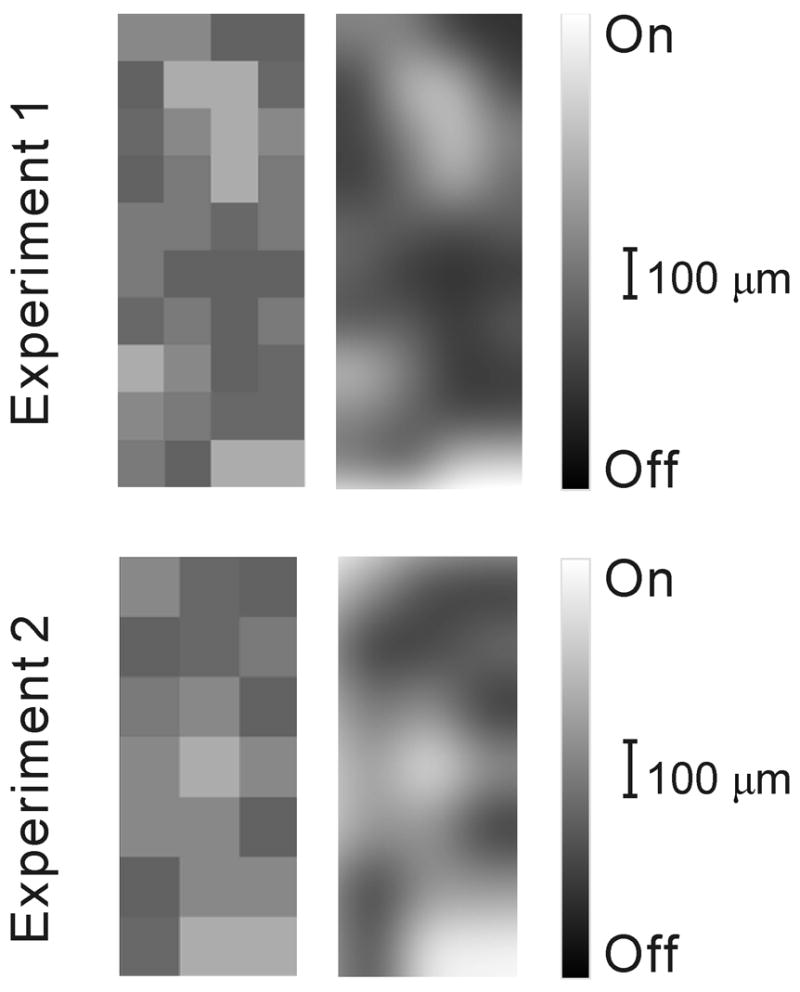
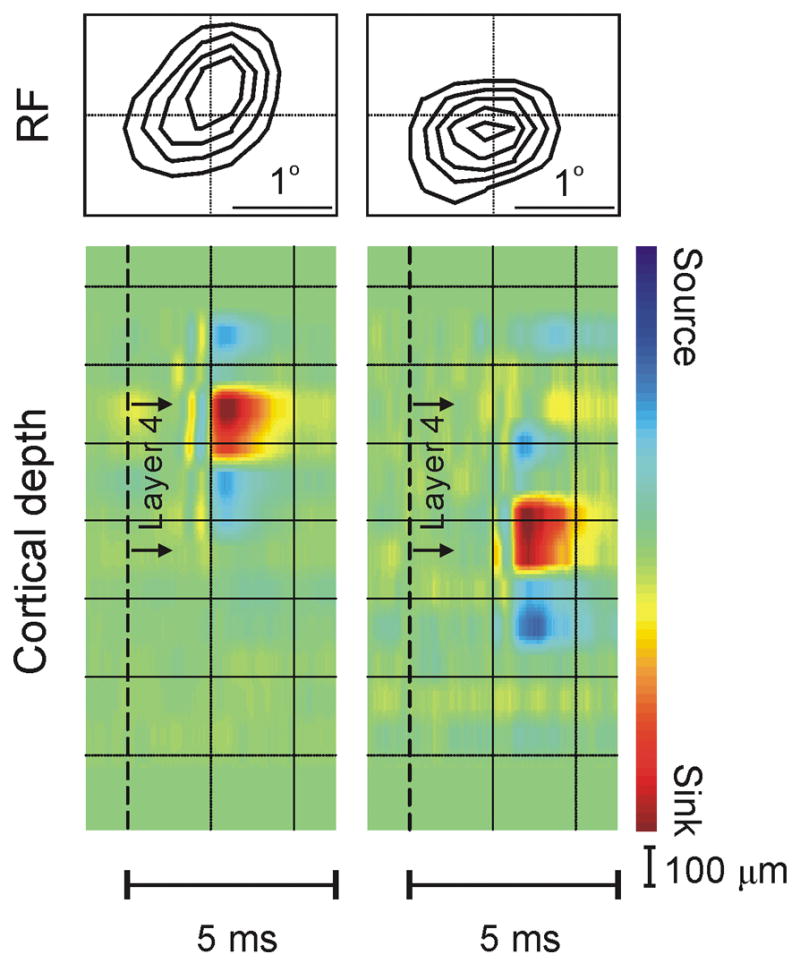
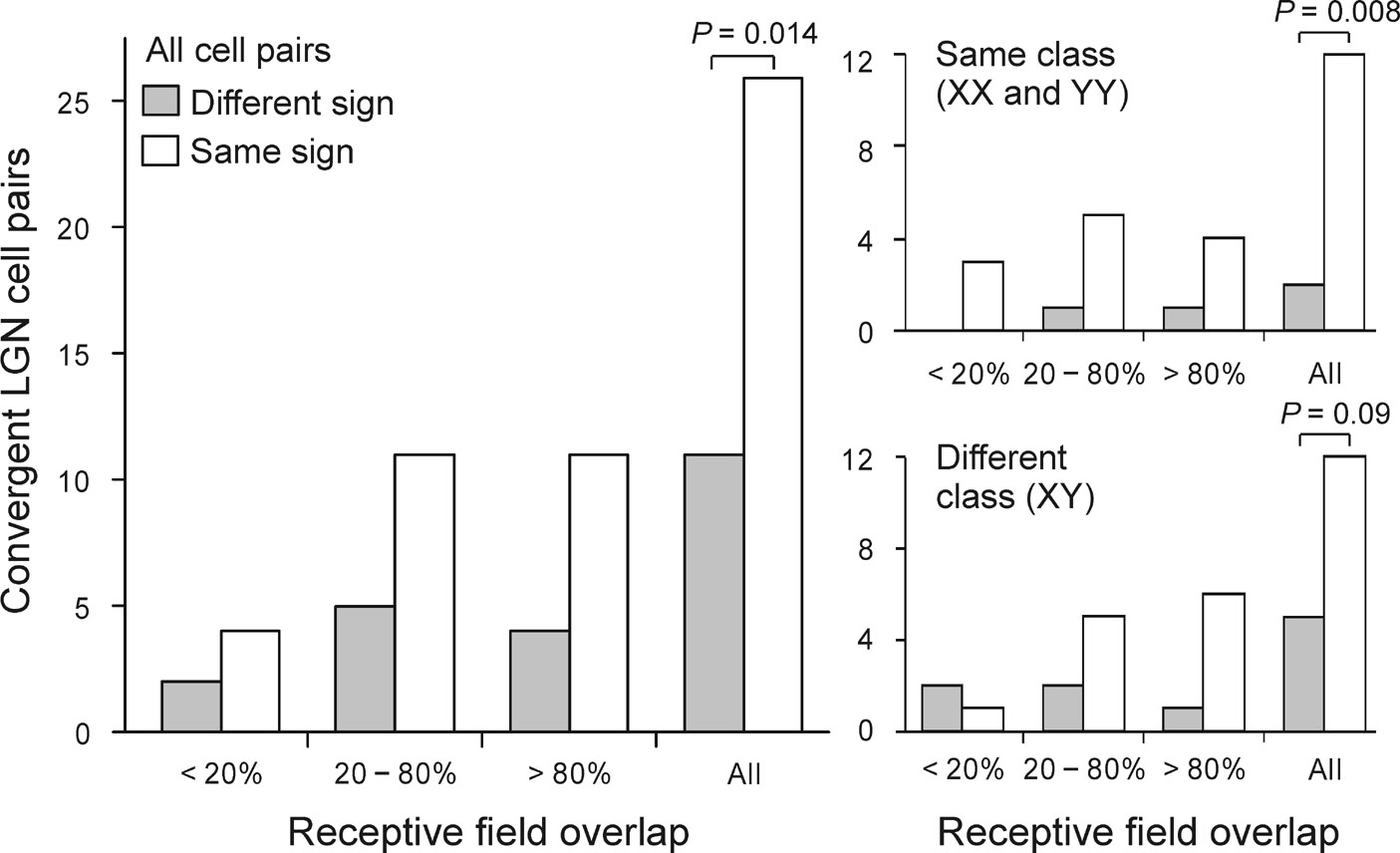
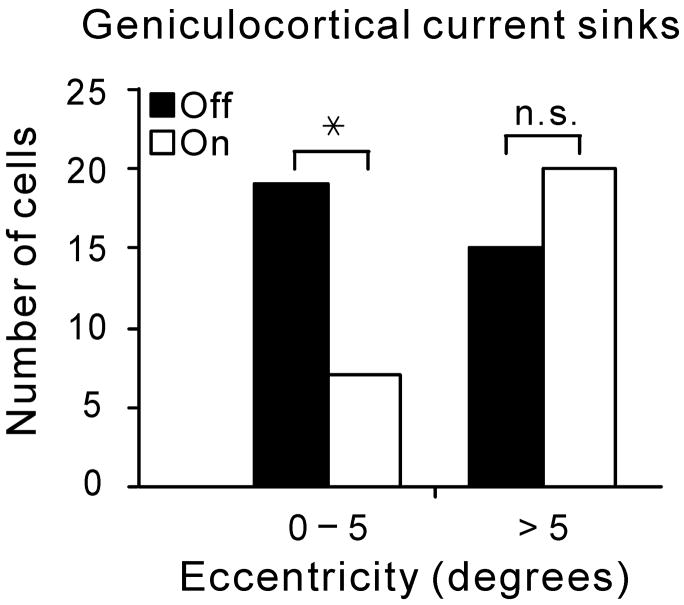
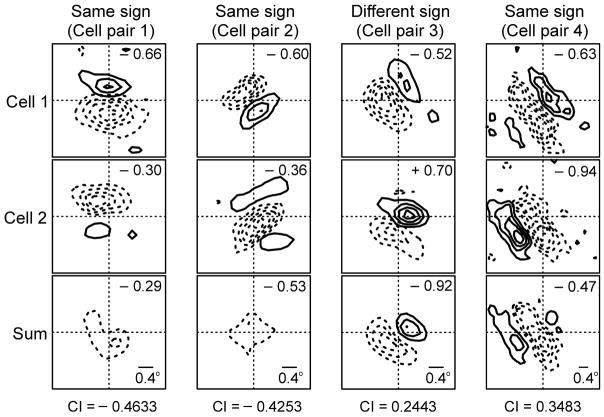
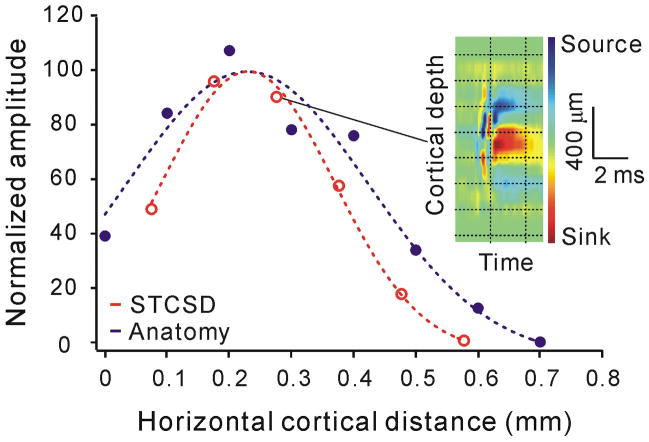
On- and off-center geniculate afferents form two major channels of visual processing that are thought to converge in the primary visual cortex. However, humans with severely reduced on responses can have normal visual acuity when tested in a white background, which indicates that off channels can function relatively independently from on channels under certain conditions. Consistent with this functional independence of channels, we demonstrate here that on- and off-center geniculate afferents segregate in different domains of the cat primary visual cortex and that off responses dominate the cortical representation of the area centralis. On average, 70% of the geniculate afferents converging at the same cortical domain had receptive fields of the same contrast polarity. Moreover, off-center afferents dominated the representation of the area centralis in the cortex, but not in the thalamus, indicating that on- and off-center afferents are balanced in number, but not in the amount of cortical territory that they cover.
Figures







References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
