

Physical linkage of naturally complexed bacterial outer membrane proteins enhances immunogenicity
- PMID: 18086812
- PMCID: PMC2258817
- DOI: 10.1128/IAI.01356-07
Physical linkage of naturally complexed bacterial outer membrane proteins enhances immunogenicity
Abstract
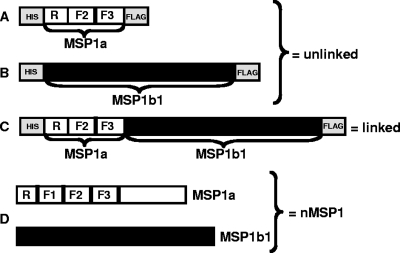
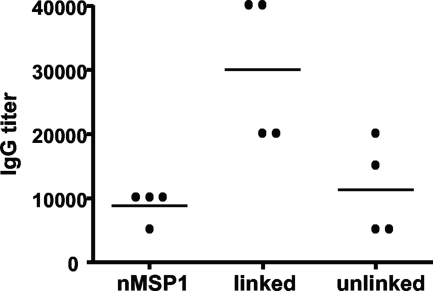
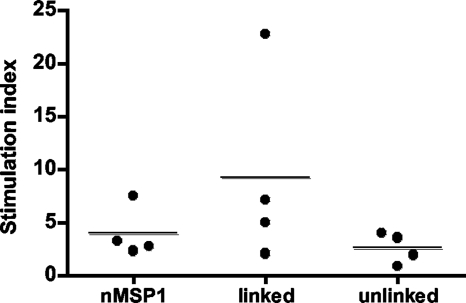
The outer membrane proteins (OMPs) of bacterial pathogens are essential for their growth and survival and especially for attachment and invasion of host cells. Since the outer membrane is the interface between the bacterium and the host cell, outer membranes and individual OMPs are targeted for development of vaccines against many bacterial diseases. Whole outer membrane fractions often protect against disease, and this protection cannot be fully reproduced by using individual OMPs. Exactly how the interactions among individual OMPs influence immunity is not well understood. We hypothesized that one OMP rich in T-cell epitopes can act as a carrier for an associated OMP which is poor in T-cell epitopes to generate T-dependent antibody responses, similar to the hapten-carrier effect. Major surface protein 1a (MSP1a) and MSP1b1 occur as naturally complexed OMPs in the Anaplasma marginale outer membrane. Previous studies demonstrated that immunization with the native MSP1 heteromer induced strong immunoglobulin G (IgG) responses to both proteins, but only MSP1a stimulated strong CD4+ T-cell responses. Therefore, to test our hypothesis, constructs of CD4+ T-cell epitopes from MSP1a linked to MSP1b1 were compared with individually administered MSP1a and MSP1b1 for induction of MSP1b-specific IgG. By linking the T-cell epitopes from MSP1a to MSP1b1, significantly higher IgG titers against MSP1b1 were induced. Understanding how the naturally occurring intermolecular interactions between OMPs influence the immune response may lead to more effective vaccine design.
Figures
References
-
- Abbott, J. R., G. H. Palmer, K. A. Kegerreis, P. F. Hetrick, C. J. Howard, J. C. Hope, and W. C. Brown. 2005. Rapid and long-term disappearance of CD4+ T lymphocyte responses specific for Anaplasma marginale major surface protein-2 (MSP2) in MSP2 vaccinates following challenge with live A. marginale. J. Immunol. 1746702-6715. - PubMed
-
- Ben-Yedidia, T., and R. Arnon. 2006. Flagella as a platform for epitope-based vaccines. Isr. Med. Assoc. J. 8316-318. - PubMed
-
- Ben-Yedidia, T., and R. Arnon. 2005. Towards an epitope-based human vaccine for influenza. Hum. Vaccines 195-101. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
