

The mitochondrion is a site of trypanocidal action of the aromatic diamidine DB75 in bloodstream forms of Trypanosoma brucei
- PMID: 18086841
- PMCID: PMC2258549
- DOI: 10.1128/AAC.00642-07
The mitochondrion is a site of trypanocidal action of the aromatic diamidine DB75 in bloodstream forms of Trypanosoma brucei
Abstract
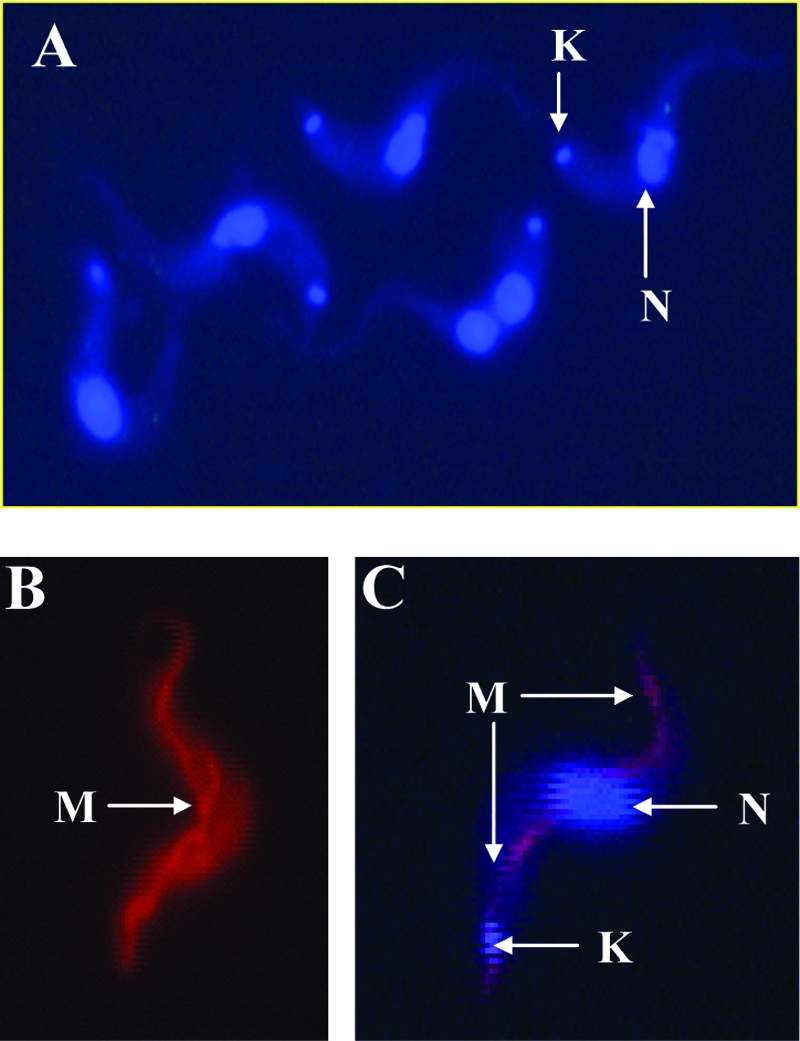
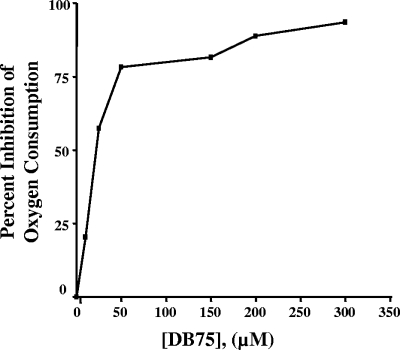
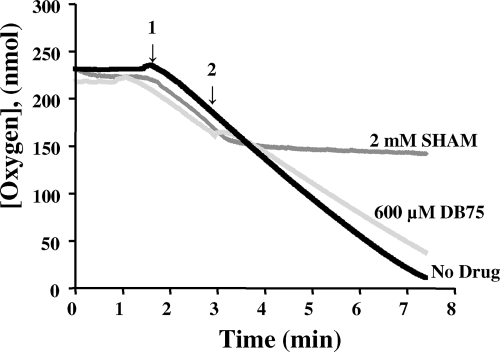
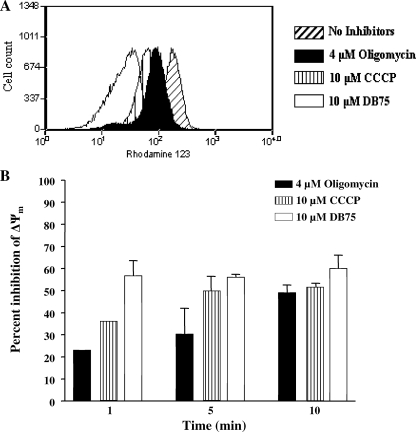
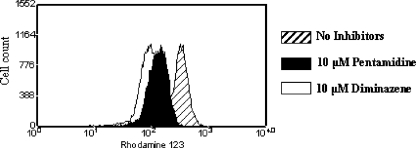
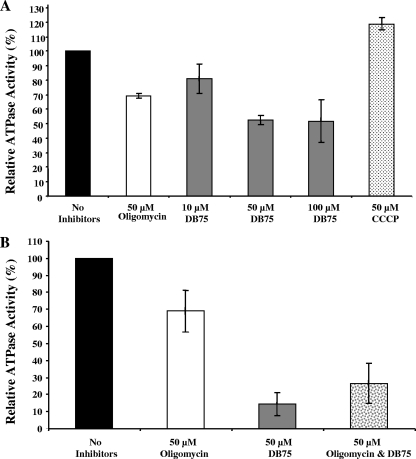
Human African trypanosomiasis (HAT) is a fatal tropical disease caused by infection with protozoans of the species Trypanosoma brucei gambiense and T. b. rhodesiense. An oral prodrug, DB289, is a promising new therapy undergoing phase III clinical trials for early-stage HAT. DB289 is metabolically converted to the active trypanocidal diamidine DB75 [2,5-bis(4-amidinophenyl)furan]. We previously determined that DB75 inhibits yeast mitochondrial function (C. A. Lanteri, B. L. Trumpower, R. R. Tidwell, and S. R. Meshnick, Antimicrob. Agent Chemother. 48:3968-3974, 2004). The purpose of this study was to investigate if DB75 targets the mitochondrion of T. b. brucei bloodstream forms. DB75 rapidly accumulates within the mitochondria of living trypanosomes, as indicated by the fluorescent colocalization of DB75 with a mitochondrion-specific dye. Fluorescence-activated cell sorting analysis of rhodamine 123-stained living trypanosomes shows that DB75 and other trypanocidal diamidines (pentamidine and diminazene) collapse the mitochondrial membrane potential. DB75 inhibits ATP hydrolysis within T. brucei mitochondria and appears to inhibit the oligomycin-sensitive F 1 F 0-ATPase and perhaps other ATPases. DB75 is most likely not an inhibitor of electron transport within trypanosome mitochondria, since DB75 fails to inhibit mitochondrial respiration when glycerol-3-phosphate is used as the respiratory substrate. However, DB75 inhibits whole-cell respiration (50% inhibitory concentration, 20 microM) at drug concentrations and incubation durations that also result in the dissipation of the mitochondrial membrane potential. Taken together, these findings suggest that the mitochondrion is a target of the trypanocidal action of DB75.
Figures
References
-
- Allemann, N., and A. Schneider. 2000. ATP production in isolated mitochondria of procyclic Trypanosoma brucei. Mol. Biochem. Parasitol. 111:87-94. - PubMed
-
- Barrett, M. P., R. J. Burchmore, A. Stich, J. O. Lazzari, A. C. Frasch, J. J. Cazzulo, and S. Krishna. 2003. The trypanosomiases. Lancet 362:1469-1480. - PubMed
-
- Bienen, E. J., G. C. Hill, and K. O. Shin. 1983. Elaboration of mitochondrial function during Trypanosoma brucei differentiation. Mol. Biochem. Parasitol. 7:75-86. - PubMed
-
- Bienen, E. J., R. K. Maturi, G. Pollakis, and A. B. Clarkson, Jr. 1993. Non-cytochrome mediated mitochondrial ATP production in bloodstream form Trypanosoma brucei brucei. Eur. J. Biochem. 216:75-80. - PubMed
-
- Bouteille, B., O. Oukem, S. Bisser, and M. Dumas. 2003. Treatment perspectives for human African trypanosomiasis. Fundam. Clin. Pharmacol. 17:171-181. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
