

The dermis contains langerin+ dendritic cells that develop and function independently of epidermal Langerhans cells
- PMID: 18086861
- PMCID: PMC2150992
- DOI: 10.1084/jem.20071724
The dermis contains langerin+ dendritic cells that develop and function independently of epidermal Langerhans cells
Abstract
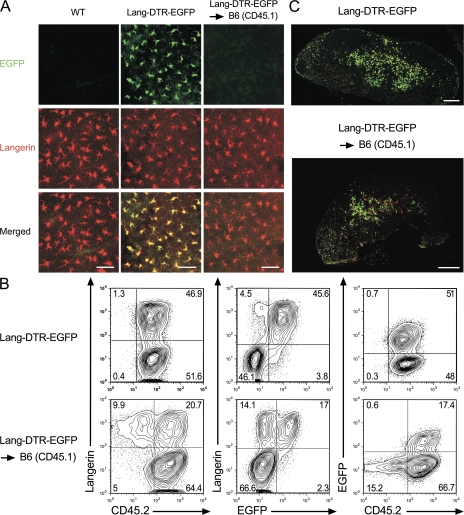
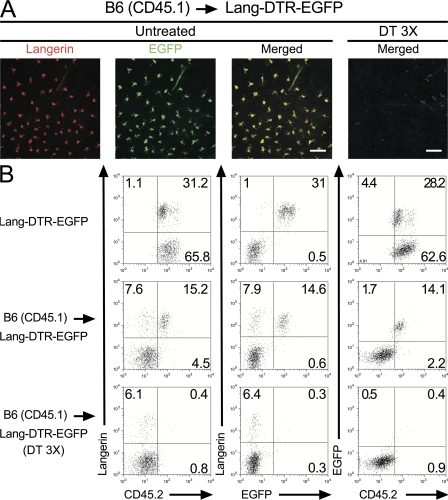
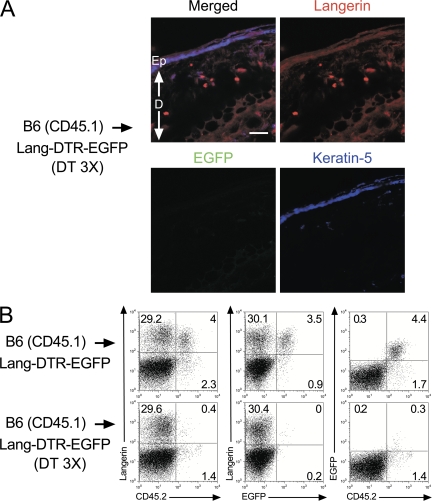
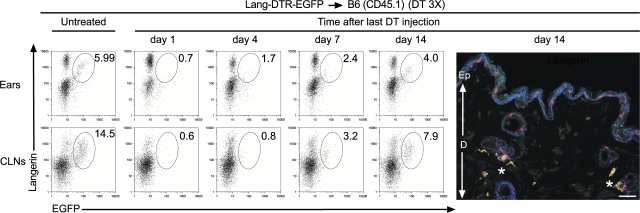
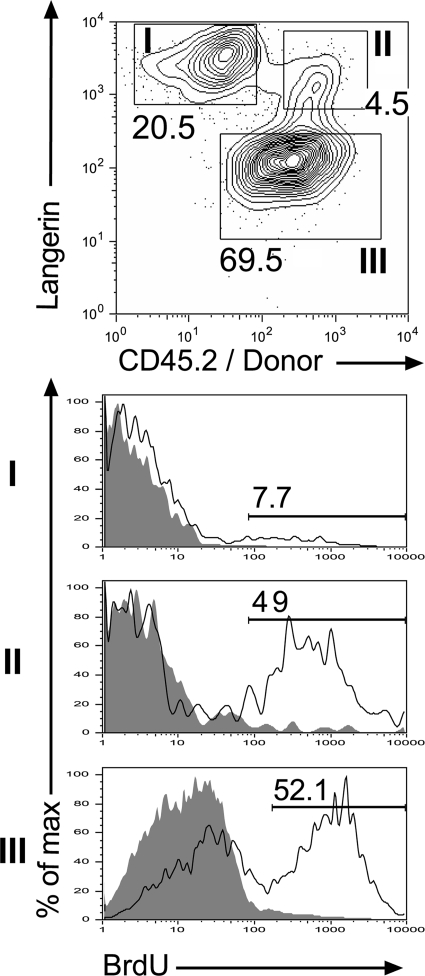
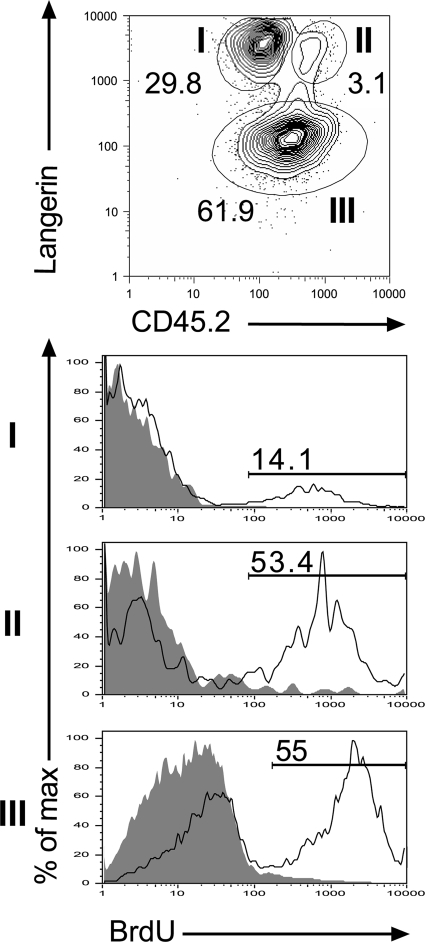
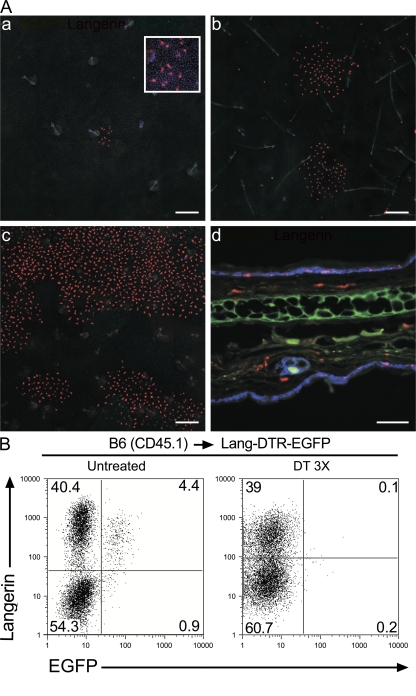
Langerhans cells (LCs) constitute a subset of dendritic cells (DCs) that express the lectin langerin and that reside in their immature state in epidermis. Paradoxically, in mice permitting diphtheria toxin (DT)-mediated ablation of LCs, epidermal LCs reappeared with kinetics that lagged behind that of their putative progeny found in lymph nodes (LNs). Using bone marrow (BM) chimeras, we showed that a major fraction of the langerin(+), skin-derived DCs found in LNs originates from a developmental pathway that is independent from that of epidermal LCs. This pathway, the existence of which was unexpected, originates in the dermis and gives rise to langerin(+) dermal DCs (DDCs) that should not be confused with epidermal LCs en route to LNs. It explains that after DT treatment, some langerin(+), skin-derived DCs reappear in LNs long before LC-derived DCs. Using CD45 expression and BrdU-labeling kinetics, both LCs and langerin(+) DDCs were found to coexist in wild-type mice. Moreover, DT-mediated ablation of epidermal LCs opened otherwise filled niches and permitted repopulation of adult noninflammatory epidermis with BM-derived LCs. Our results stress that the langerin(+) DC network is more complex than originally thought and have implications for the development of transcutaneous vaccines and the improvement of humanized mouse models.
Figures
References
-
- Villadangos, J.A., and P. Schnorrer. 2007. Intrinsic and cooperative antigen-presenting functions of dendritic-cell subsets in vivo. Nat. Rev. Immunol. 7:543–555. - PubMed
-
- Larregina, A.T., and L.D. Falo. 2005. Changing paradigms in cutaneous immunology: adapting with dendritic cells. J. Invest. Dermatol. 124:1–12. - PubMed
-
- Carbone, F.R., G.T. Belz, and W.R. Heath. 2004. Transfer of antigen between migrating and lymph node-resident DCs in peripheral T-cell tolerance and immunity. Trends Immunol. 25:655–658. - PubMed
-
- Shortman, K., and S.H. Naik. 2007. Steady-state and inflammatory dendritic-cell development. Nat. Rev. Immunol. 7:19–30. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
