

Two RNA polymerase I subunits control the binding and release of Rrn3 during transcription
- PMID: 18086878
- PMCID: PMC2258765
- DOI: 10.1128/MCB.01464-07
Two RNA polymerase I subunits control the binding and release of Rrn3 during transcription
Abstract
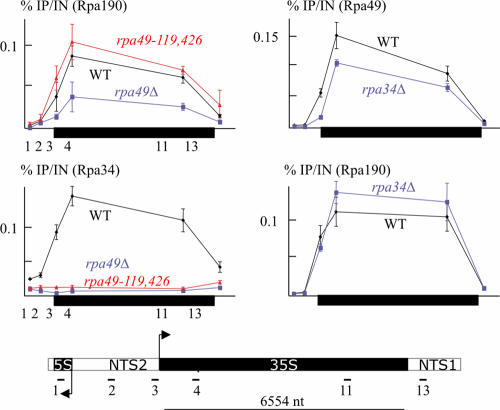
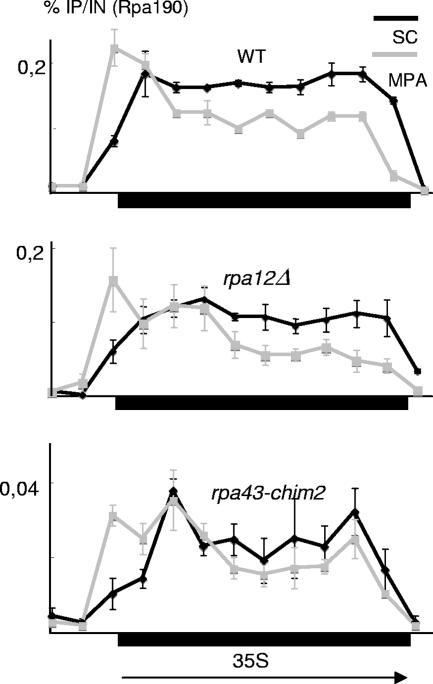
Rpa34 and Rpa49 are nonessential subunits of RNA polymerase I, conserved in species from Saccharomyces cerevisiae and Schizosaccharomyces pombe to humans. Rpa34 bound an N-terminal region of Rpa49 in a two-hybrid assay and was lost from RNA polymerase in an rpa49 mutant lacking this Rpa34-binding domain, whereas rpa34Delta weakened the binding of Rpa49 to RNA polymerase. rpa34Delta mutants were caffeine sensitive, and the rpa34Delta mutation was lethal in a top1Delta mutant and in rpa14Delta, rpa135(L656P), and rpa135(D395N) RNA polymerase mutants. These defects were shared by rpa49Delta mutants, were suppressed by the overexpression of Rpa49, and thus, were presumably mediated by Rpa49 itself. rpa49 mutants lacking the Rpa34-binding domain behaved essentially like rpa34Delta mutants, but strains carrying rpa49Delta and rpa49-338::HIS3 (encoding a form of Rpa49 lacking the conserved C terminus) had reduced polymerase occupancy at 30 degrees C, failed to grow at 25 degrees C, and were sensitive to 6-azauracil and mycophenolate. Mycophenolate almost fully dissociated the mutant polymerase from its ribosomal DNA (rDNA) template. The rpa49Delta and rpa49-338::HIS3 mutations had a dual effect on the transcription initiation factor Rrn3 (TIF-IA). They partially impaired its recruitment to the rDNA promoter, an effect that was bypassed by an N-terminal deletion of the Rpa43 subunit encoded by rpa43-35,326, and they strongly reduced the release of the Rrn3 initiation factor during elongation. These data suggest a dual role of the Rpa49-Rpa34 dimer during the recruitment of Rrn3 and its subsequent dissociation from the elongating polymerase.
Figures







References
-
- Aslett, M., and V. Wood. 2006. Gene Ontology annotation status of the fission yeast genome: preliminary coverage approaches 100%. Yeast 23913-919. - PubMed
-
- Bell, S. P., R. M. Learned, H. M. Jantzen, and R. Tjian. 1988. Functional cooperativity between transcription factors UBF1 and SL1 mediates ribosomal RNA synthesis. Science 2411192-1197. - PubMed
-
- Bier, M., S. Fath, and H. Tschochner. 2004. The composition of the RNA polymerase I transcription machinery switches from initiation to elongation mode. FEBS Lett. 56441-46. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases