

The newly synthesized pool of dopamine determines the severity of methamphetamine-induced neurotoxicity
- PMID: 18088364
- PMCID: PMC2668123
- DOI: 10.1111/j.1471-4159.2007.05155.x
The newly synthesized pool of dopamine determines the severity of methamphetamine-induced neurotoxicity
Abstract
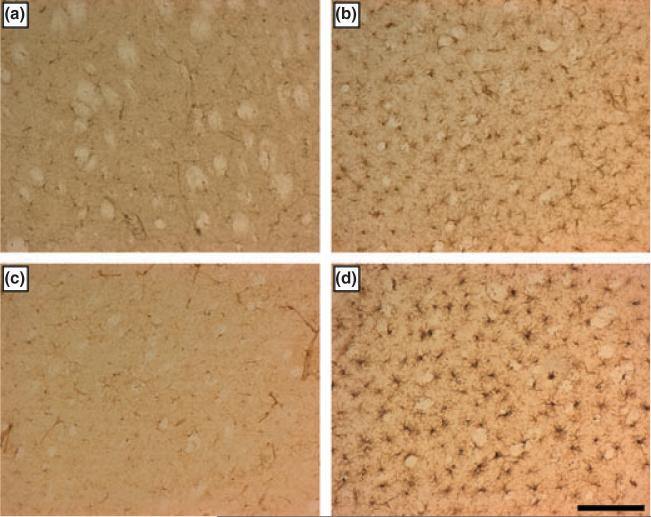
The neurotransmitter dopamine (DA) has long been implicated as a participant in the neurotoxicity caused by methamphetamine (METH), yet, its mechanism of action in this regard is not fully understood. Treatment of mice with the tyrosine hydroxylase (TH) inhibitor alpha-methyl-p-tyrosine (AMPT) lowers striatal cytoplasmic DA content by 55% and completely protects against METH-induced damage to DA nerve terminals. Reserpine, by disrupting vesicle amine storage, depletes striatal DA by more than 95% and accentuates METH-induced neurotoxicity. l-DOPA reverses the protective effect of AMPT against METH and enhances neurotoxicity in animals with intact TH. Inhibition of MAO-A by clorgyline increases pre-synaptic DA content and enhances METH striatal neurotoxicity. In all conditions of altered pre-synaptic DA homeostasis, increases or decreases in METH neurotoxicity paralleled changes in striatal microglial activation. Mice treated with AMPT, l-DOPA, or clorgyline + METH developed hyperthermia to the same extent as animals treated with METH alone, whereas mice treated with reserpine + METH were hypothermic, suggesting that the effects of alterations in cytoplasmic DA on METH neurotoxicity were not strictly mediated by changes in core body temperature. Taken together, the present data reinforce the notion that METH-induced release of DA from the newly synthesized pool of transmitter into the extracellular space plays an essential role in drug-induced striatal neurotoxicity and microglial activation. Subtle alterations in intracellular DA content can lead to significant enhancement of METH neurotoxicity. Our results also suggest that reactants derived from METH-induced oxidation of released DA may serve as neuronal signals that lead to microglial activation early in the neurotoxic process associated with METH.
Figures







References
-
- Albers DS, Sonsalla PK. Methamphetamine-induced hyperthermia and dopaminergic neurotoxicity in mice: pharmacological profile of protective and nonprotective agents. J. Pharmacol. Exp. Ther. 1995;275:1104–1114. - PubMed
-
- Benkovic SA, O'Callaghan JP, Miller DB. Sensitive indicators of injury reveal hippocampal damage in C57BL/6J mice treated with kainic acid in the absence of tonic-clonic seizures. Brain Res. 2004;1024:59–76. - PubMed
-
- Block ML, Zecca L, Hong JS. Microglia-mediated neurotoxicity: uncovering the molecular mechanisms. Nat Rev Neurosci. 2007;8:57–69. - PubMed
-
- Bowyer JF, Davies DL, Schmued L, Broening HW, Newport GD, Slikker W, Jr, Holson RR. Further studies of the role of hyperthermia in methamphetamine neurotoxicity. J. Pharmacol. Exp. Ther. 1994;268:1571–1580. - PubMed
-
- Brown IR. Induction of heat shock (stress) genes in the mammalian brain by hyperthermia and other traumatic events: a current perspective. J. Neurosci. Res. 1990;27:247–255. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
