

Structural analysis of the synaptic protein neuroligin and its beta-neurexin complex: determinants for folding and cell adhesion
- PMID: 18093521
- PMCID: PMC2703725
- DOI: 10.1016/j.neuron.2007.11.013
Structural analysis of the synaptic protein neuroligin and its beta-neurexin complex: determinants for folding and cell adhesion
Abstract
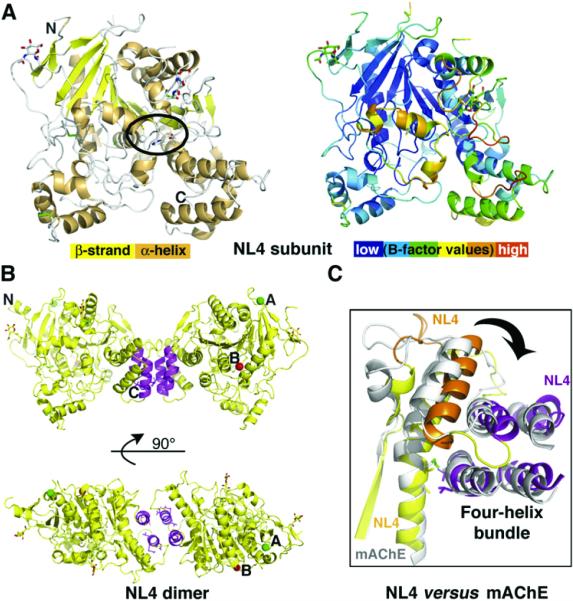
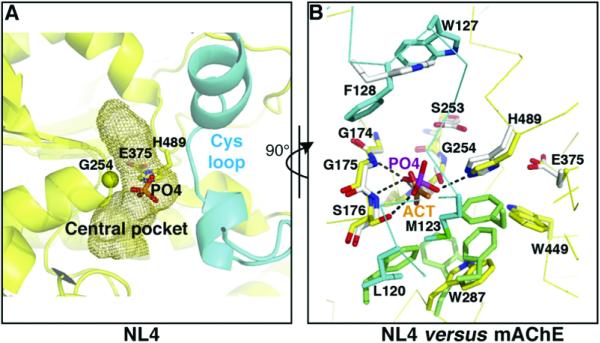
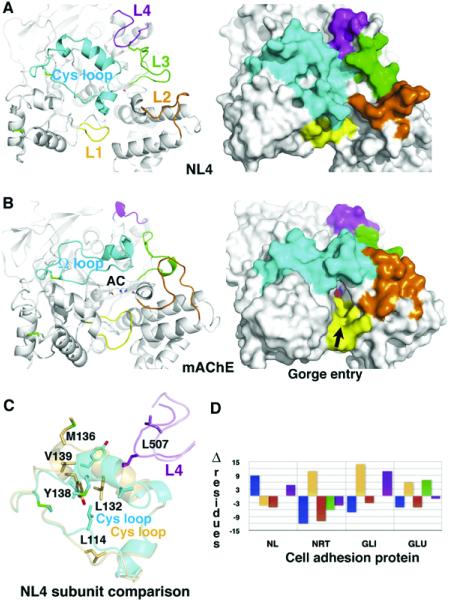
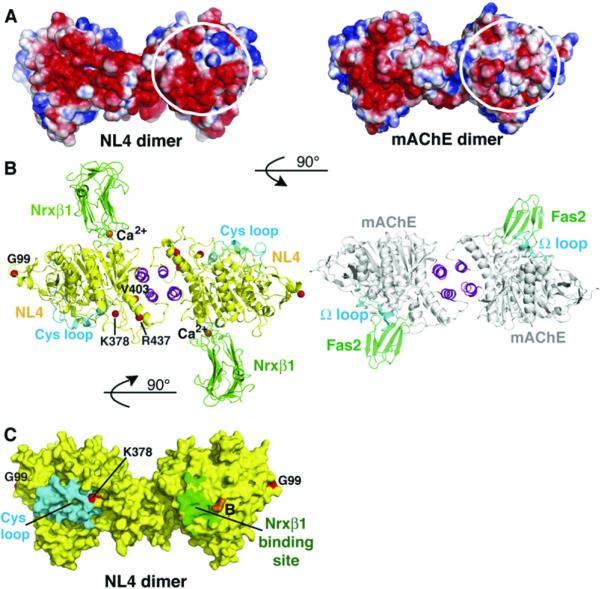
The neuroligins are postsynaptic cell adhesion proteins whose associations with presynaptic neurexins participate in synaptogenesis. Mutations in the neuroligin and neurexin genes appear to be associated with autism and mental retardation. The crystal structure of a neuroligin reveals features not found in its catalytically active relatives, such as the fully hydrophobic interface forming the functional neuroligin dimer; the conformations of surface loops surrounding the vestigial active center; the location of determinants that are critical for folding and processing; and the absence of a macromolecular dipole and presence of an electronegative, hydrophilic surface for neurexin binding. The structure of a beta-neurexin-neuroligin complex reveals the precise orientation of the bound neurexin and, despite a limited resolution, provides substantial information on the Ca2+-dependent interactions network involved in trans-synaptic neurexin-neuroligin association. These structures exemplify how an alpha/beta-hydrolase fold varies in surface topography to confer adhesion properties and provide templates for analyzing abnormal processing or recognition events associated with autism.
Figures

Comment in
-
A crystal-clear interaction: relating neuroligin/neurexin complex structure to function at the synapse.Neuron. 2007 Dec 20;56(6):937-9. doi: 10.1016/j.neuron.2007.12.003. Neuron. 2007. PMID: 18093514
Comment on
-
Structures of neuroligin-1 and the neuroligin-1/neurexin-1 beta complex reveal specific protein-protein and protein-Ca2+ interactions.Neuron. 2007 Dec 20;56(6):992-1003. doi: 10.1016/j.neuron.2007.12.002. Neuron. 2007. PMID: 18093522
References
-
- Auld VJ, Fetter RD, Broadie K, Goodman CS. Gliotactin, a novel transmembrane protein on peripheral glia, is required to form the blood-nerve barrier in Drosophila. Cell. 1995;81:757–767. - PubMed
-
- Blasi F, Bacchelli E, Pesaresi G, Carone S, Bailey AJ, Maestrini E. Absence of coding mutations in the X-linked genes neuroligin 3 and neuroligin 4 in individuals with autism from the IMGSAC collection. Am. J. Med. Genet. B. Neuropsychiatr. Genet. 2006;141:220–221. - PubMed
-
- Botti SA, Felder CE, Sussman JL, Silman I. Electro-tactins: a class of adhesion proteins with conserved electrostatic and structural motifs. Protein Eng. 1998;11:415–420. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
