

Interleukin-10 expression in macrophages during phagocytosis of apoptotic cells is mediated by homeodomain proteins Pbx1 and Prep-1
- PMID: 18093541
- PMCID: PMC2194654
- DOI: 10.1016/j.immuni.2007.11.014
Interleukin-10 expression in macrophages during phagocytosis of apoptotic cells is mediated by homeodomain proteins Pbx1 and Prep-1
Abstract
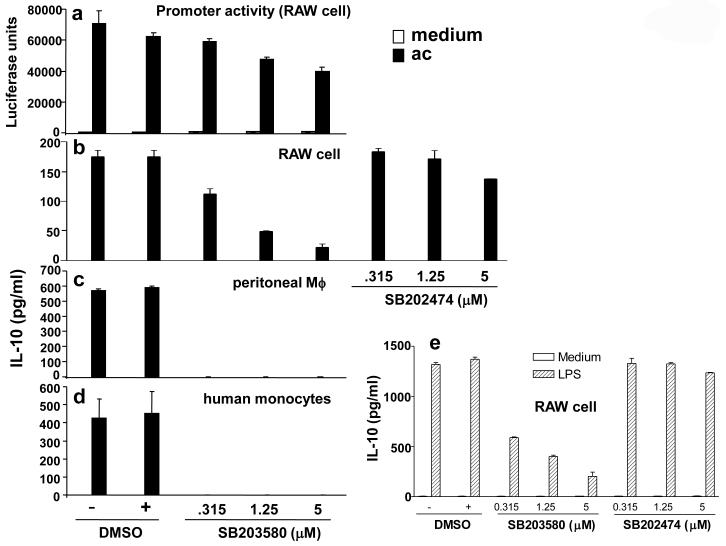
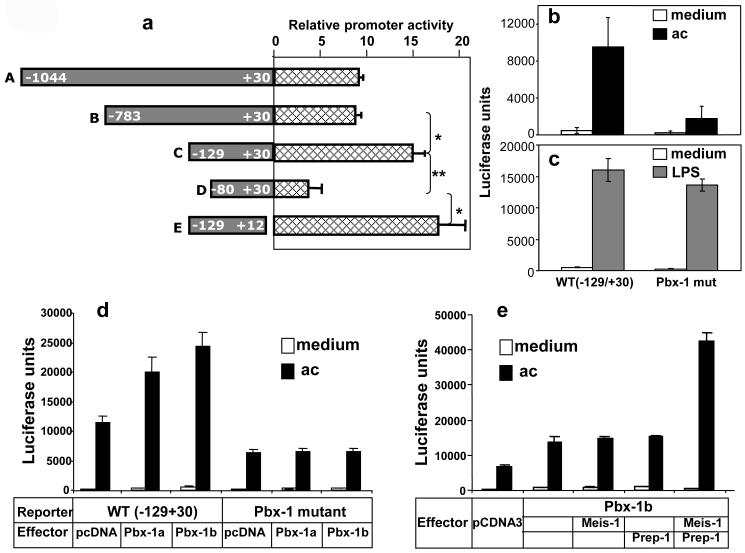
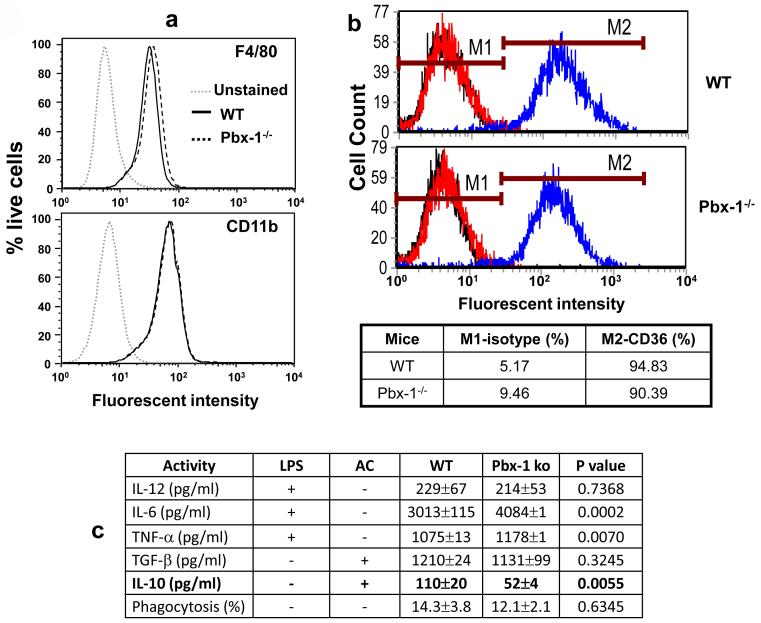
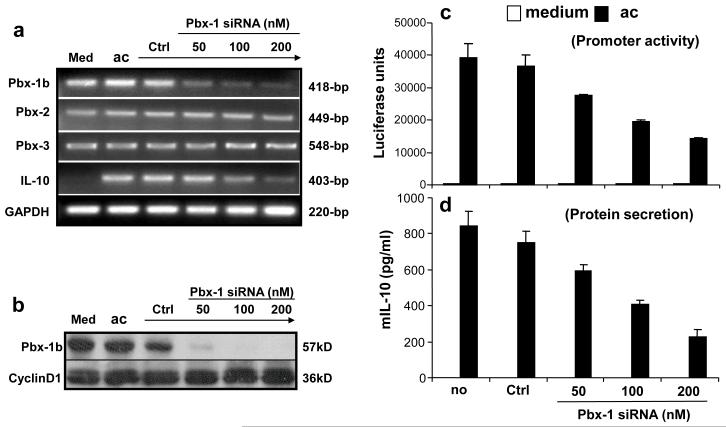
Production of interleukin (IL)-10, a major immunoregulatory cytokine, by phagocytes during clearance of apoptotic cells is critical to ensuring cellular homeostasis and suppression of autoimmunity. Little is known about the regulatory mechanisms in this fundamental process. We report that IL-10 production stimulated by apoptotic cells was regulated at the point of transcription in a manner dependent on p38 mitogen-activated protein kinase, partially on the scavenger receptor CD36, and required cell-cell contact but not phagocytosis. By using a reporter assay, we mapped the apoptotic-cell-response element (ACRE) in the human IL10 promoter and provide biochemical and physiological evidence that ACRE mediates the transcriptional activation of IL10 by pre-B cell leukemia transcription factor-1b and another Hox cofactor Pbx-regulating protein 1 in response to apoptotic cells. This study establishes a role of two developmentally critical factors (Pbx1 and Prep-1) in the regulation of homeostasis in the immune system.
Figures



References
-
- Aderem A, Underhill DM. Mechanisms of phagocytosis in macrophages. Annu Rev Immunol. 1999;17:593–623. - PubMed
-
- Benkhart EM, Siedlar M, Wedel A, Werner T, Ziegler-Heitbrock HW. Role of Stat3 in lipopolysaccharide-induced IL-10 gene expression. J Immunol. 2000a;165:1612–1617. - PubMed
-
- Benkhart EM, Siedlar M, Wedel A, Werner T, Ziegler-Heitbrock HW. Role of Stat3 in lipopolysaccharide-induced IL-10 gene expression. J Immunol. 2000b;165:1612–1617. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
