

EPLIN mediates linkage of the cadherin catenin complex to F-actin and stabilizes the circumferential actin belt
- PMID: 18093941
- PMCID: PMC2224173
- DOI: 10.1073/pnas.0710504105
EPLIN mediates linkage of the cadherin catenin complex to F-actin and stabilizes the circumferential actin belt
Abstract
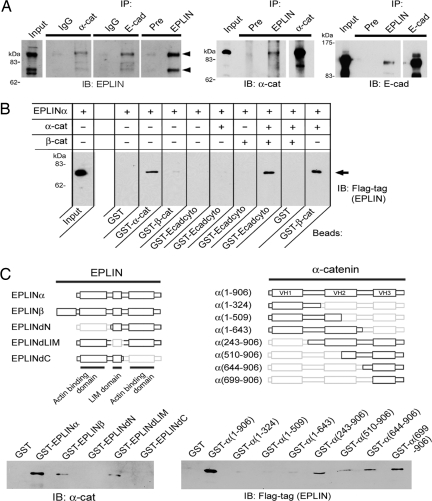
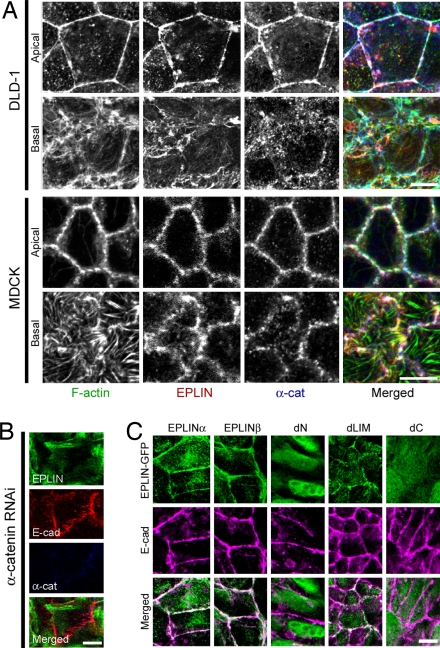
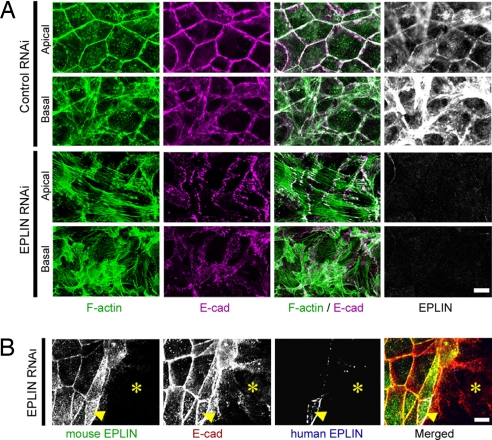
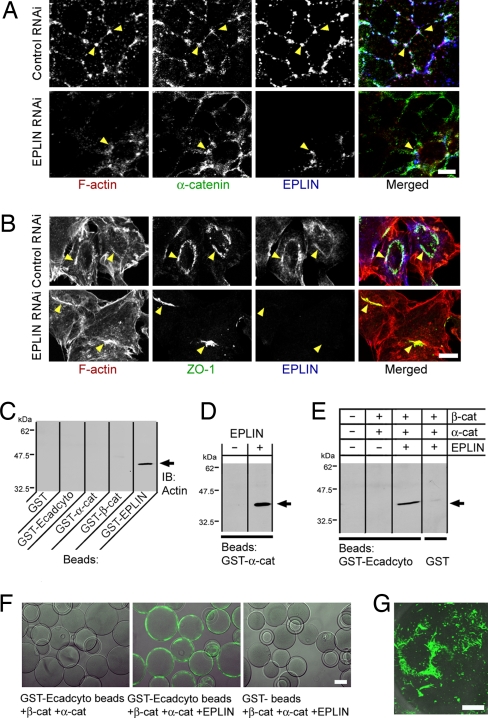
The cadherin-catenin complex is the major machinery for cell-cell adhesion in many animal species. This complex in general associates with actin fibers at its cytoplasmic side, organizing the adherens junction (AJ). In epithelial cells, the AJ encircles the cells near their apical surface and forms the "zonula adherens" or "adhesion belt." The mechanism as to how the cadherin-catenin complex and F-actin cooperate to generate these junctional structures, however, remains unknown. Here, we show that EPLIN (epithelial protein lost in neoplasm; also known as Lima-1), an actin-binding protein, couples with alpha-catenin and, in turn, links the cadherin-catenin complex to F-actin. Without EPLIN, this linkage was unable to form. When EPLIN had been depleted in epithelial cells, the adhesion belt was disorganized and converted into zipper-like junctions in which actin fibers were radially arranged. However, nonjunctional actin fibers were not particularly affected by EPLIN depletion. As EPLIN is known to have the ability to suppress actin depolymerization, our results suggest that EPLIN functions to link the cadherin-catenin complex to F-actin and simultaneously stabilizes this population of actin fibers, resulting in the establishment of the adhesion belt.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Mege RM, Gavard J, Lambert M. Curr Opin Cell Biol. 2006;18:541–548. - PubMed
-
- Lecuit T. Trends Cell Biol. 2005;15:34–42. - PubMed
-
- Pokutta S, Weis WI. Annu Rev Cell Dev Biol. 2007;23:237–261. - PubMed
-
- Hildebrand JD. J Cell Sci. 2005;118:5191–5203. - PubMed
-
- Haigo SL, Hildebrand JD, Harland RM, Wallingford JB. Curr Biol. 2003;13:2125–2137. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
