

Embryological origin of airway smooth muscle
- PMID: 18094078
- PMCID: PMC2645301
- DOI: 10.1513/pats.200704-049VS
Embryological origin of airway smooth muscle
Abstract
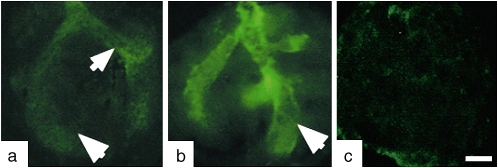
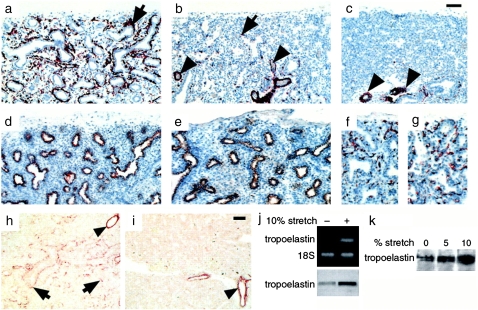
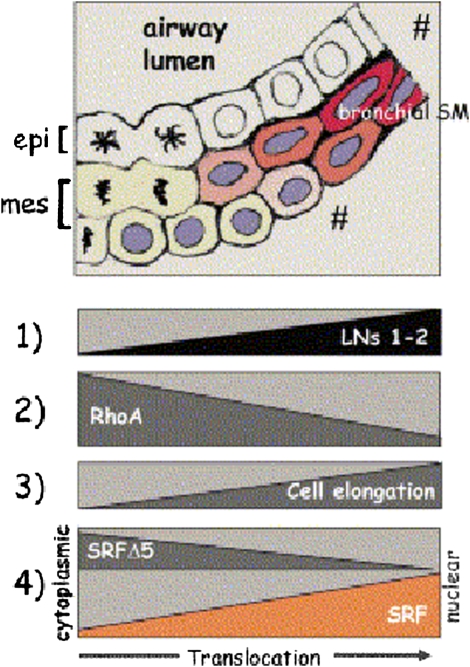
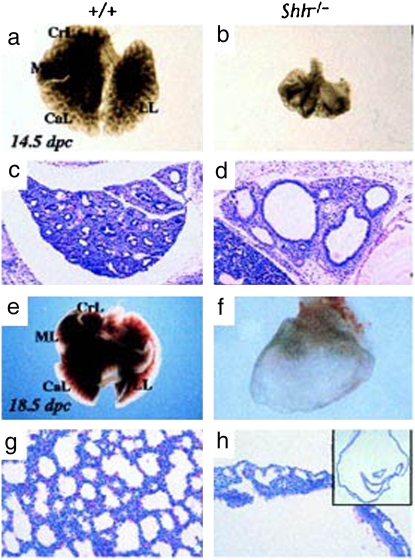
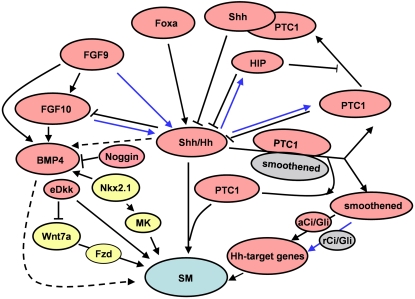
Airway smooth muscle (SM) develops from local mesenchymal cells located around the tips of growing epithelial buds. These cells gradually displace from distal to proximal position alongside the bronchial tree, elongate, and begin to synthesize SM-specific proteins. Mechanical tension (either generated by cell spreading/elongation or stretch), as well as epithelial paracrine factors, regulates the process of bronchial myogenesis. The specific roles of many of these paracrine factors during normal lung development are currently unknown. It is also unknown how and if mechanical and paracrine signals integrate into a common myogenic pathway. Furthermore, as with vascular SM and other types of visceral SM, we are just beginning to elucidate the intracellular signaling pathways and the genetic program that controls lung myogenesis. Here we present what we have learned so far about the embryogenesis of bronchial muscle.
Figures
References
-
- Sparrow MP, Lamb JP. Ontogeny of airway smooth muscle: structure, innervation, myogenesis and function in the fetal lung. Respir Physiolo Neurobiol 2003;137:361–372. - PubMed
-
- Roman J, McDonald JA. Expression of fibronectin, the integrin alpha 5, and alpha-smooth muscle actin in heart and lung development. Am J Respir Cell Mol Biol 1992;6:472–480. - PubMed
-
- McHugh KM. Molecular analysis of smooth muscle development in the mouse. Dev Dyn 1995;204:278–290. - PubMed
-
- Li L, Miano JM, Cserjesi P, Olson EN. Sm22 alpha, a marker of adult smooth muscle, is expressed in multiple myogenic lineages during embryogenesis. Circ Res 1996;78:188–195. - PubMed
-
- Mitchell HW, Sparrow MP, Tagliaferri RP. Inhibitory and excitatory responses to field stimulation in fetal and adult pig airway. Pediatr Res 1990;28:69–74. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources