

Dominant-negative FADD rescues the in vivo fitness of a cytomegalovirus lacking an antiapoptotic viral gene
- PMID: 18094168
- PMCID: PMC2258921
- DOI: 10.1128/JVI.01803-07
Dominant-negative FADD rescues the in vivo fitness of a cytomegalovirus lacking an antiapoptotic viral gene
Abstract
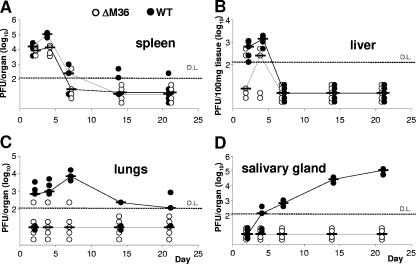
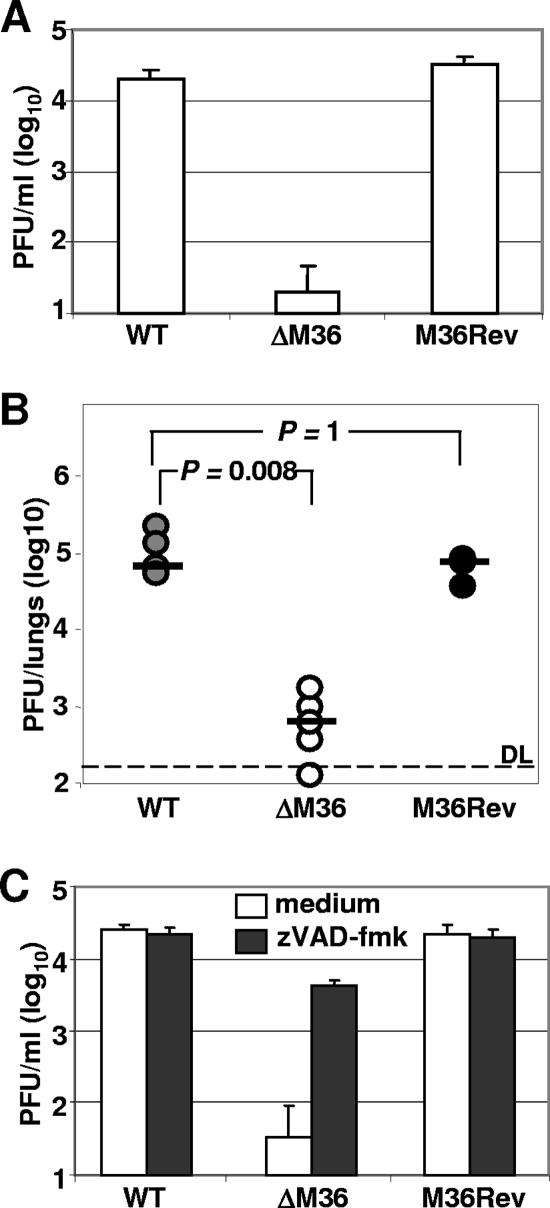
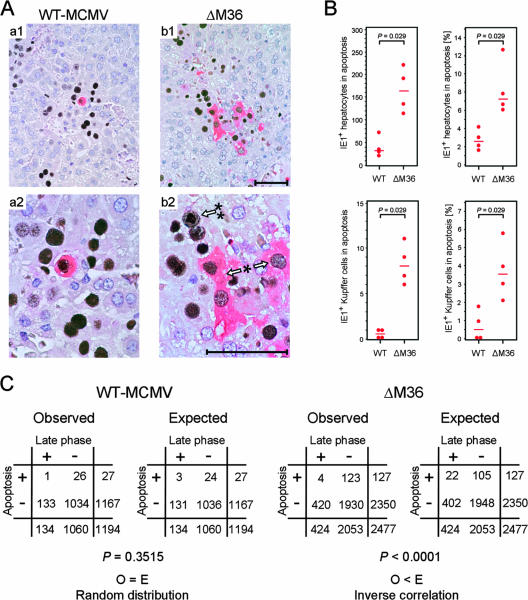
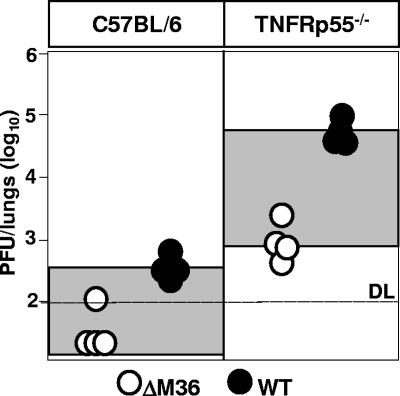
Genes that inhibit apoptosis have been described for many DNA viruses. Herpesviruses often contain even more than one gene to control cell death. Apoptosis inhibition by viral genes is postulated to contribute to viral fitness, although a formal proof is pending. To address this question, we studied the mouse cytomegalovirus (MCMV) protein M36, which binds to caspase-8 and blocks death receptor-induced apoptosis. The growth of MCMV recombinants lacking M36 (DeltaM36) was attenuated in vitro and in vivo. In vitro, caspase inhibition by zVAD-fmk blocked apoptosis in DeltaM36-infected macrophages and rescued the growth of the mutant. In vivo, DeltaM36 infection foci in liver tissue contained significantly more apoptotic hepatocytes and Kupffer cells than did revertant virus foci, and apoptosis occurred during the early phase of virus replication prior to virion assembly. To further delineate the mode of M36 function, we replaced the M36 gene with a dominant-negative FADD (FADD(DN)) in an MCMV recombinant. FADD(DN) was expressed in cells infected with the recombinant and blocked the death-receptor pathway, replacing the antiapoptotic function of M36. Most importantly, FADD(DN) rescued DeltaM36 virus replication, both in vitro and in vivo. These findings have identified the biological role of M36 and define apoptosis inhibition as a key determinant of viral fitness.
Figures
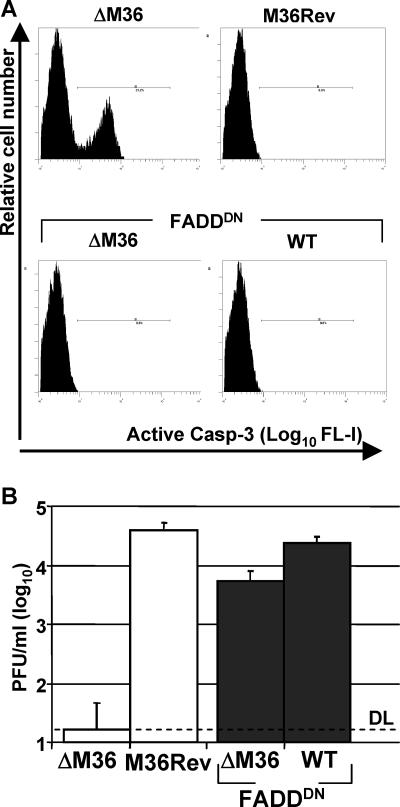
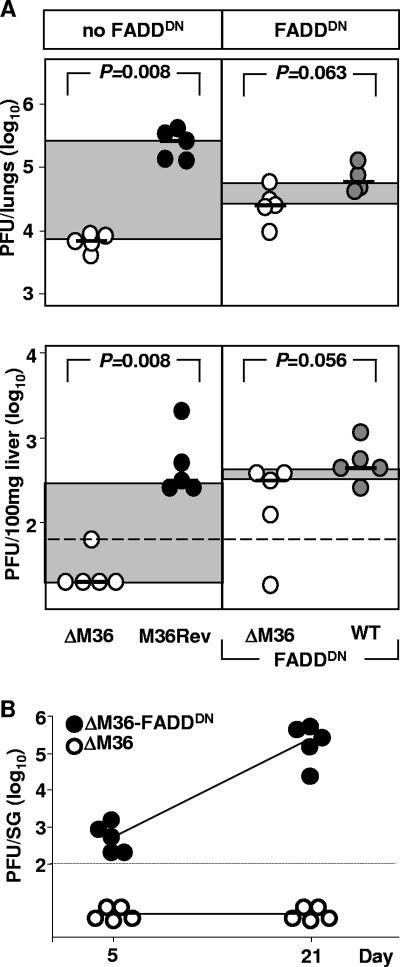
References
-
- Andoniou, C. E., and M. A. Degli-Esposti. 2006. Insights into the mechanisms of CMV-mediated interference with cellular apoptosis. Immunol. Cell Biol. 8499-106. - PubMed
-
- Askenasy, N., E. S. Yolcu, I. Yaniv, and H. Shirwan. 2005. Induction of tolerance using Fas ligand: a double-edged immunomodulator. Blood 1051396-1404. - PubMed
-
- Bertin, J., R. C. Armstrong, S. Ottilie, D. A. Martin, Y. Wang, S. Banks, G. H. Wang, T. G. Senkevich, E. S. Alnemri, B. Moss, M. J. Lenardo, K. J. Tomaselli, and J. I. Cohen. 1997. Death effector domain-containing herpesvirus and poxvirus proteins inhibit both Fas- and TNFR1-induced apoptosis. Proc. Natl. Acad. Sci. USA 941172-1176. - PMC - PubMed
-
- Brune, W., C. Menard, J. Heesemann, and U. H. Koszinowski. 2001. A ribonucleotide reductase homolog of cytomegalovirus and endothelial cell tropism. Science 291303-305. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
