

The G59S mutation in p150(glued) causes dysfunction of dynactin in mice
- PMID: 18094236
- PMCID: PMC2367233
- DOI: 10.1523/JNEUROSCI.4226-07.2007
The G59S mutation in p150(glued) causes dysfunction of dynactin in mice
Abstract
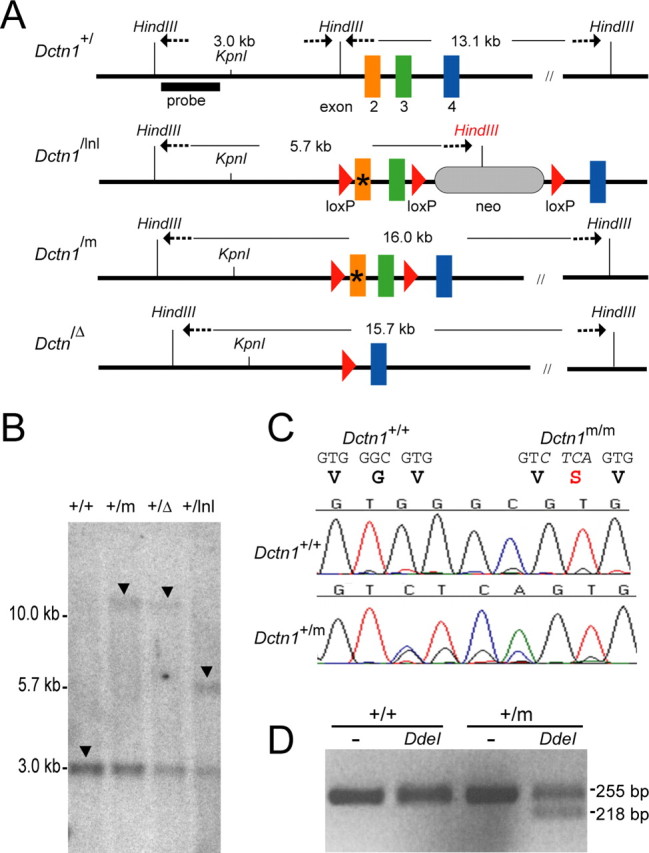
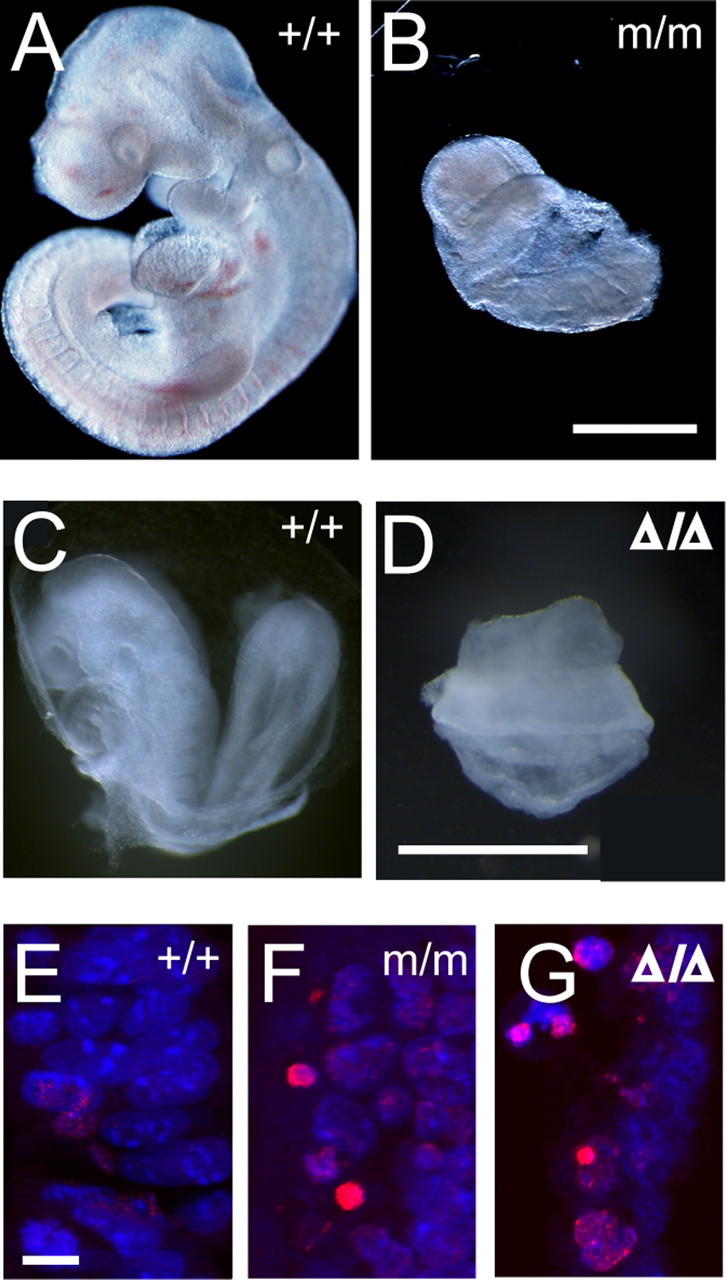
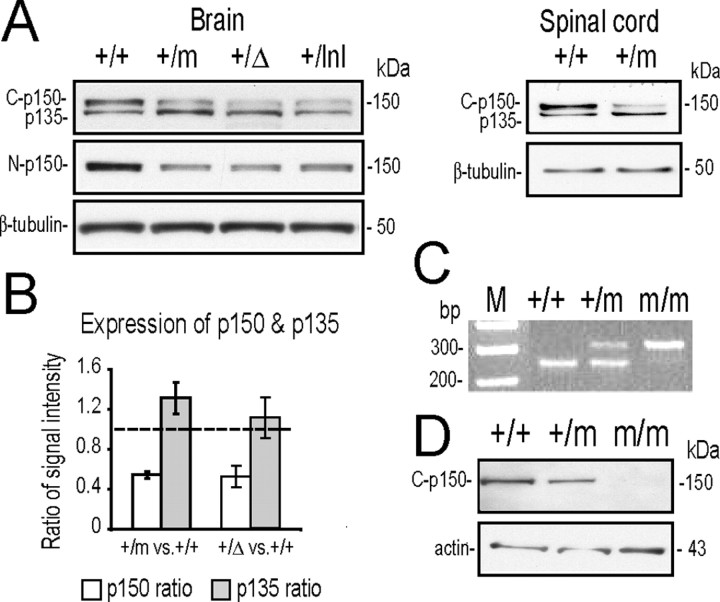
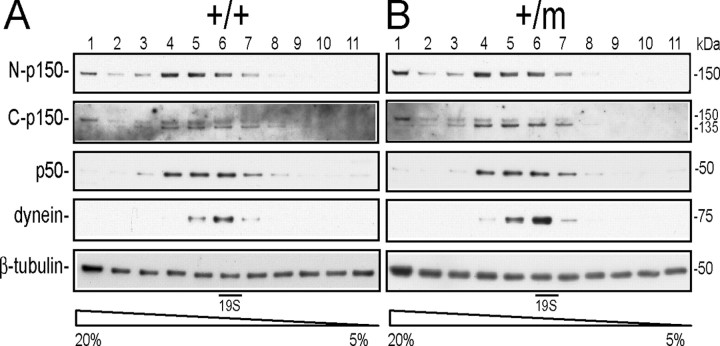
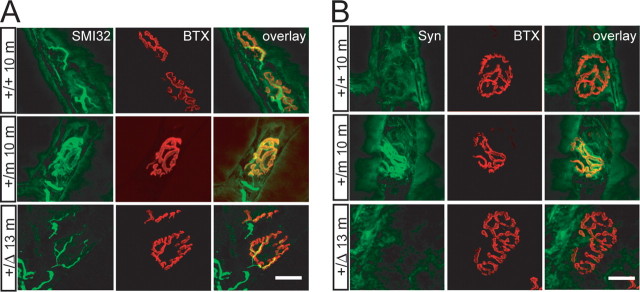
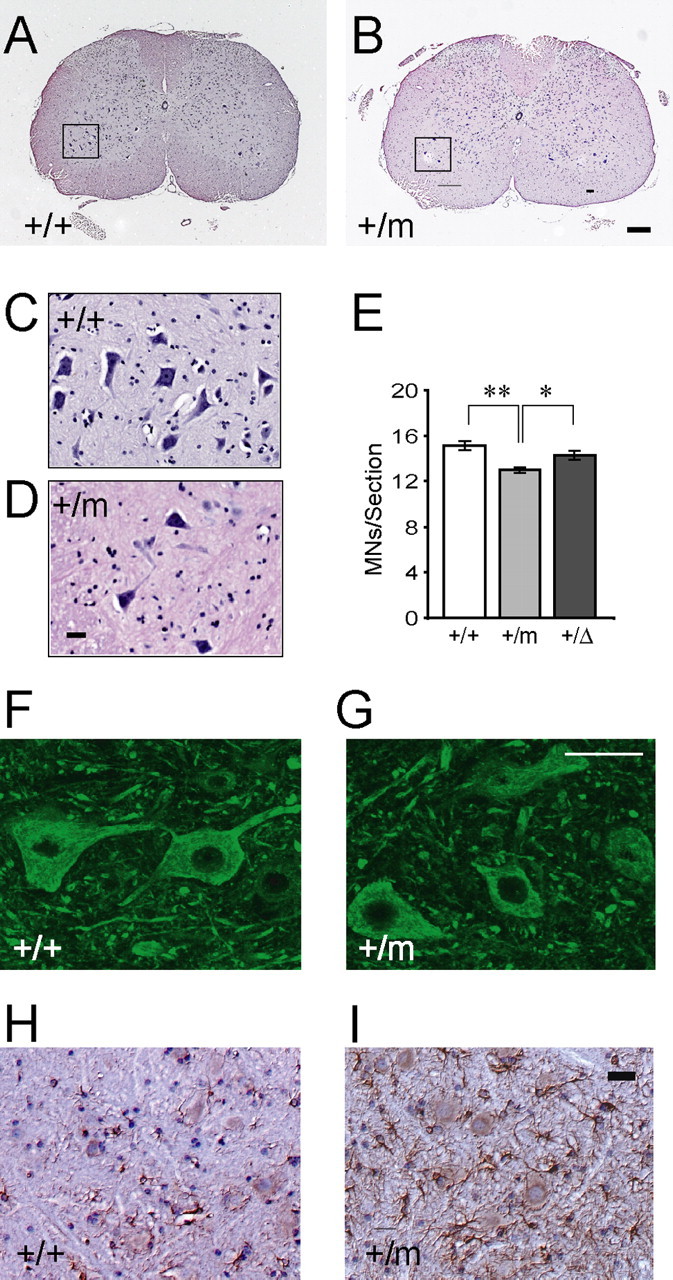
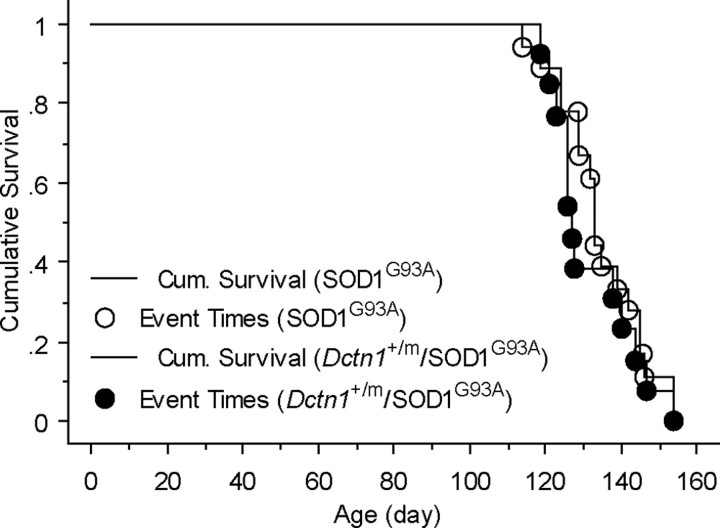
The G59S missense mutation at the conserved microtubule-binding domain of p150(glued), a major component of dynein/dynactin complex, has been linked to an autosomal dominant form of motor neuron disease (MND). To study how this mutation affects the function of the dynein/dynactin complex and contributes to motor neuron degeneration, we generated p150(glued) G59S knock-in mice. We found that the G59S mutation destabilizes p150(glued) and disrupts the function of dynein/dynactin complex, resulting in early embryonic lethality of homozygous knock-in mice. Heterozygous knock-in mice, which developed normally, displayed MND-like phenotypes after 10 months of age, including excessive accumulation of cytoskeletal and synaptic vesicle proteins at neuromuscular junctions, loss of spinal motor neurons, increase of reactive astrogliosis, and shortening of gait compared with wild-type littermates and age-matched p150(glued) heterozygous knock-out mice. Our findings indicate that the G59S mutation in p150(glued) abrogates the normal function of p150(glued) and accelerates motor neuron degeneration.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Allan V. Motor proteins: a dynamic duo. Curr Biol. 1996;6:630–633. - PubMed
-
- Bross P, Corydon TJ, Andresen BS, Jorgensen MM, Bolund L, Gregersen N. Protein misfolding and degradation in genetic diseases. Hum Mutat. 1999;14:186–198. - PubMed
-
- Culver-Hanlon TL, Lex SA, Stephens AD, Quintyne NJ, King SJ. A microtubule-binding domain in dynactin increases dynein processivity by skating along microtubules. Nat Cell Biol. 2006;8:264–270. - PubMed
-
- Eaton BA, Fetter RD, Davis GW. Dynactin is necessary for synapse stabilization. Neuron. 2002;34:729–741. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases