

Expression and localization of RGS9-2/G 5/R7BP complex in vivo is set by dynamic control of its constitutive degradation by cellular cysteine proteases
- PMID: 18094251
- PMCID: PMC6673529
- DOI: 10.1523/JNEUROSCI.3884-07.2007
Expression and localization of RGS9-2/G 5/R7BP complex in vivo is set by dynamic control of its constitutive degradation by cellular cysteine proteases
Abstract
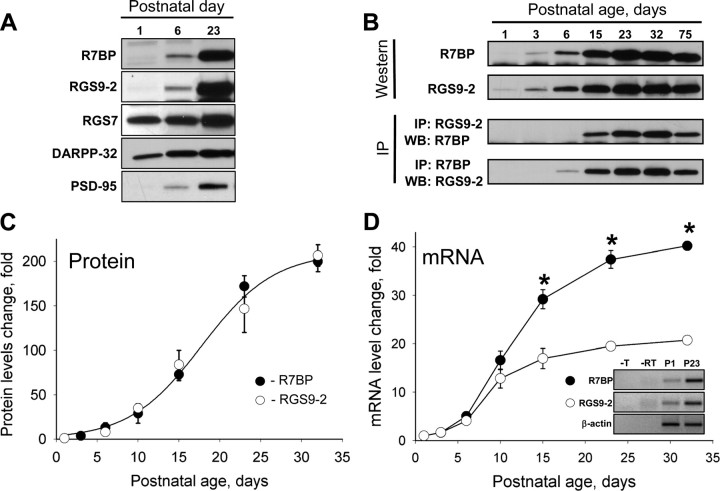
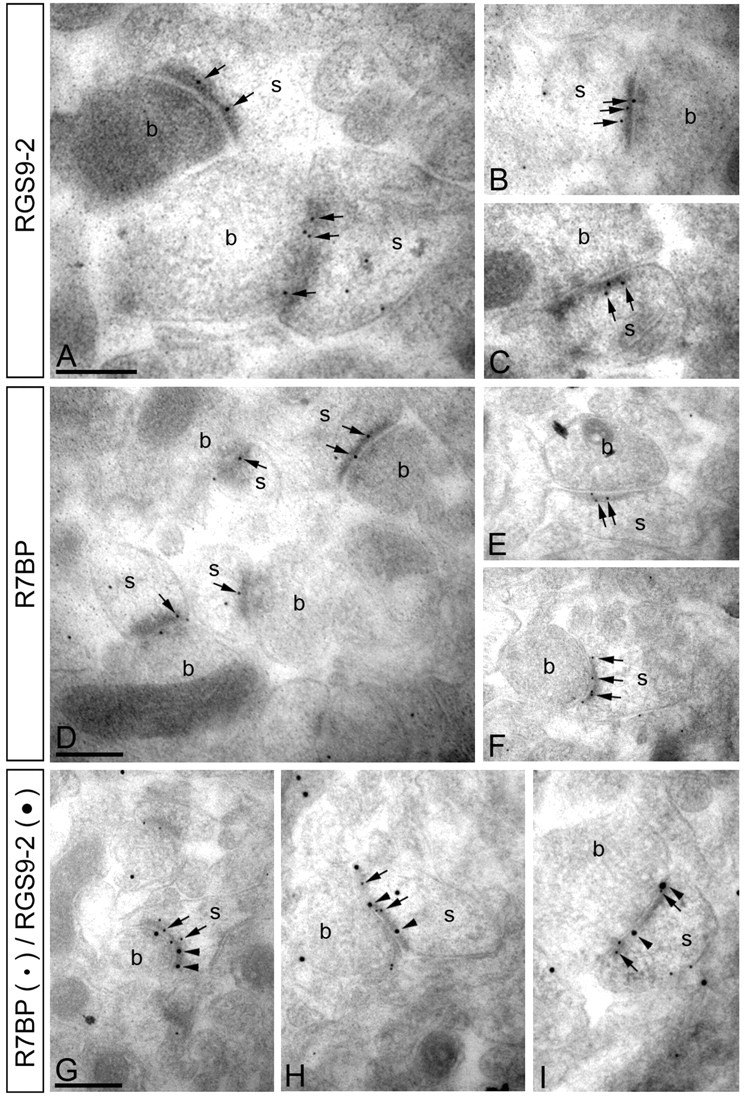
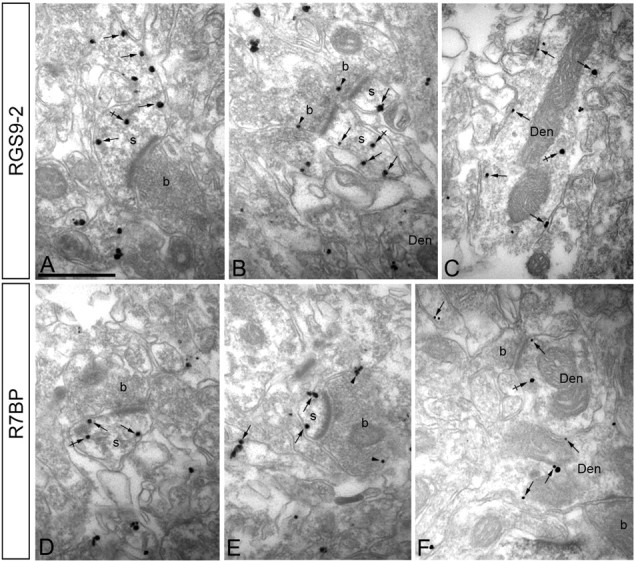
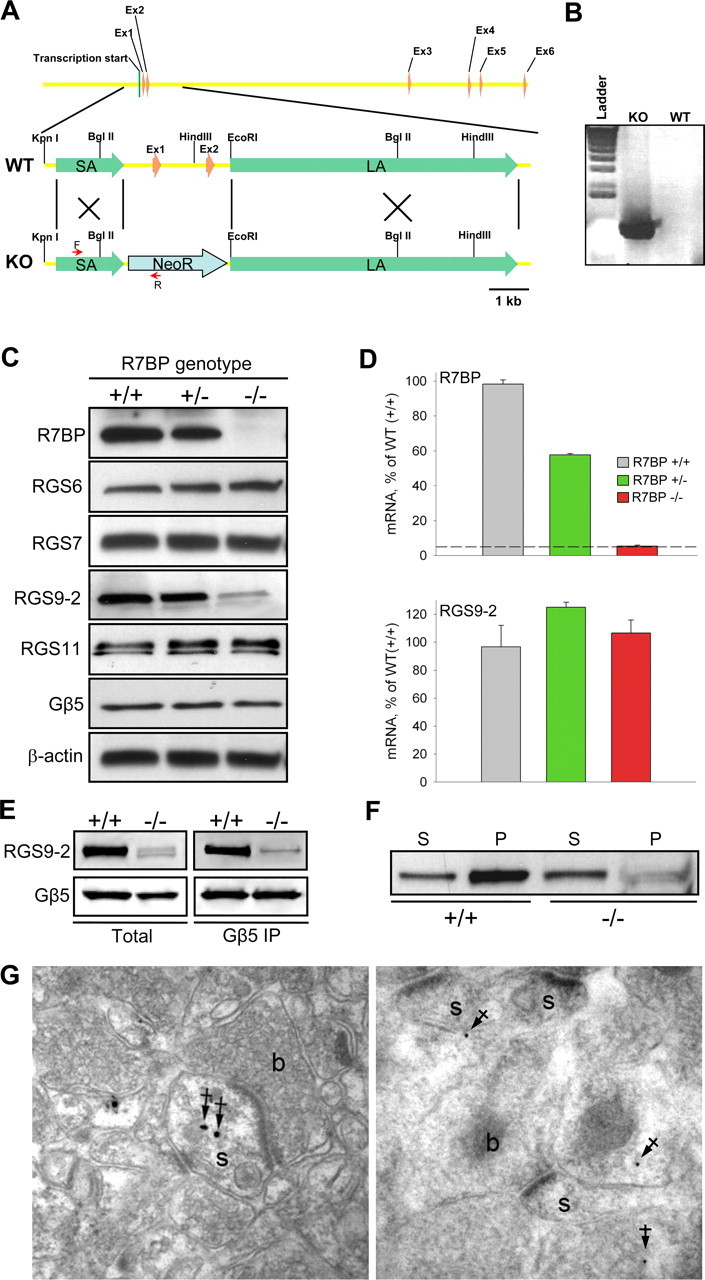
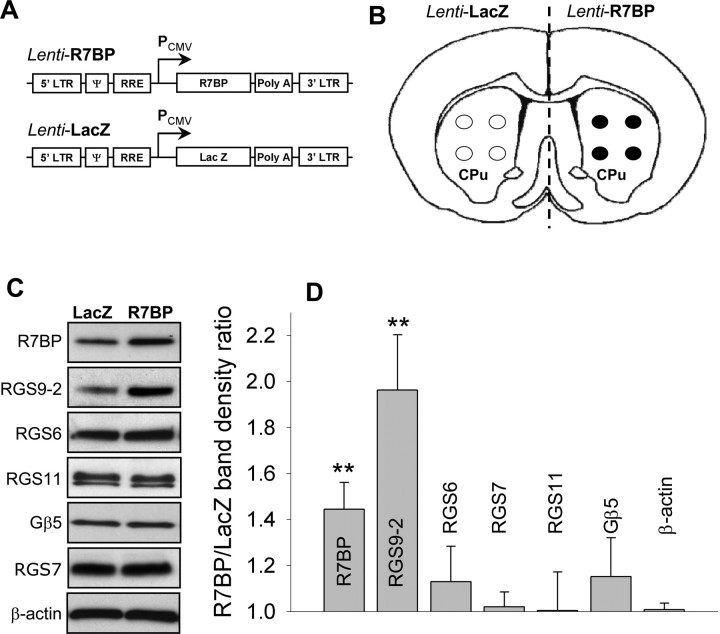
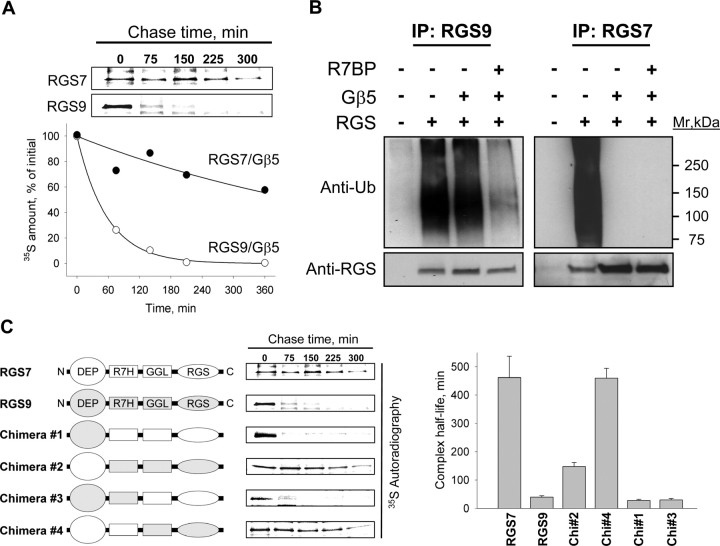
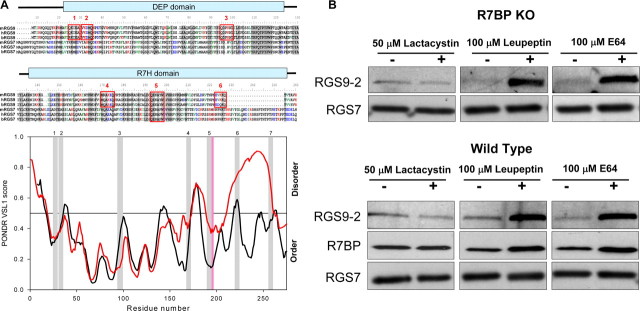
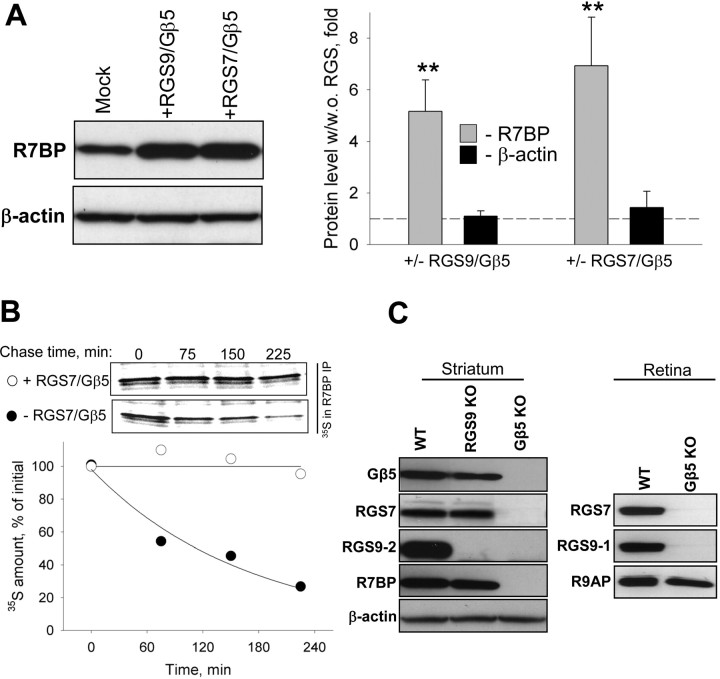
A member of regulator of G-protein signaling family, RGS9-2, is an essential modulator of signaling through neuronal dopamine and opioid G-protein-coupled receptors. Recent findings indicate that the abundance of RGS9-2 determines sensitivity of signaling in the locomotor and reward systems in the striatum. In this study we report the mechanism that sets the concentration of RGS9-2 in vivo, thus controlling G-protein signaling sensitivity in the region. We found that RGS9-2 possesses specific degradation determinants which target it for constitutive destruction by lysosomal cysteine proteases. Shielding of these determinants by the binding partner R7 binding-protein (R7BP) controls RGS9-2 expression at the posttranslational level. In addition, binding to R7BP in neurons targets RGS9-2 to the specific intracellular compartment, the postsynaptic density. Implementation of this mechanism throughout ontogenetic development ensures expression of RGS9-2/type 5 G-protein beta subunit/R7BP complexes at postsynaptic sites in unison with increased signaling demands at mature synapses.
Figures
References
-
- Anderson GR, Semenov A, Song JH, Martemyanov KA. The membrane anchor R7BP controls the proteolytic stability of the striatal specific RGS protein, RGS9-2. J Biol Chem. 2007;282:4772–4781. - PubMed
-
- Bourhis JM, Canard B, Longhi S. Predicting protein disorder and induced folding: from theoretical principles to practical applications. Curr Protein Pept Sci. 2007;8:135–149. - PubMed
-
- Burchett SA, Volk ML, Bannon MJ, Granneman JG. Regulators of G-protein signaling: rapid changes in mRNA abundance in response to amphetamine. J Neurochem. 1998;70:2216–2219. - PubMed
-
- Chen CK, Burns ME, He W, Wensel TG, Baylor DA, Simon MI. Slowed recovery of rod photoresponse in mice lacking the GTPase accelerating protein RGS9-1. Nature. 2000;403:557–560. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases