

A high quality draft consensus sequence of the genome of a heterozygous grapevine variety
- PMID: 18094749
- PMCID: PMC2147077
- DOI: 10.1371/journal.pone.0001326
A high quality draft consensus sequence of the genome of a heterozygous grapevine variety
Abstract
Background: Worldwide, grapes and their derived products have a large market. The cultivated grape species Vitis vinifera has potential to become a model for fruit trees genetics. Like many plant species, it is highly heterozygous, which is an additional challenge to modern whole genome shotgun sequencing. In this paper a high quality draft genome sequence of a cultivated clone of V. vinifera Pinot Noir is presented.
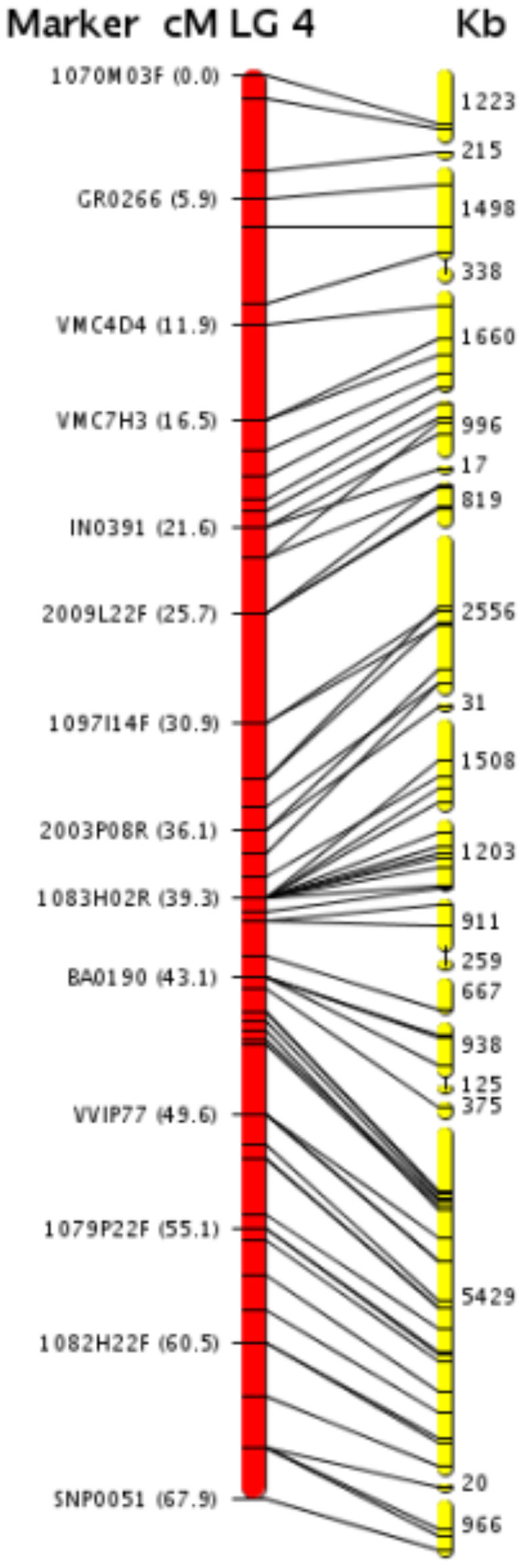
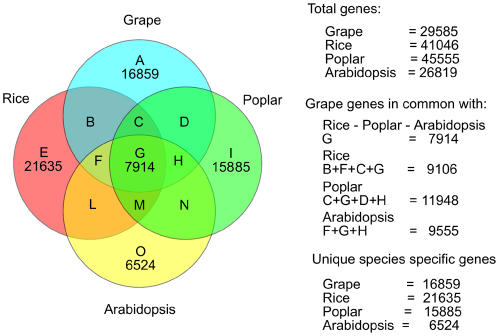
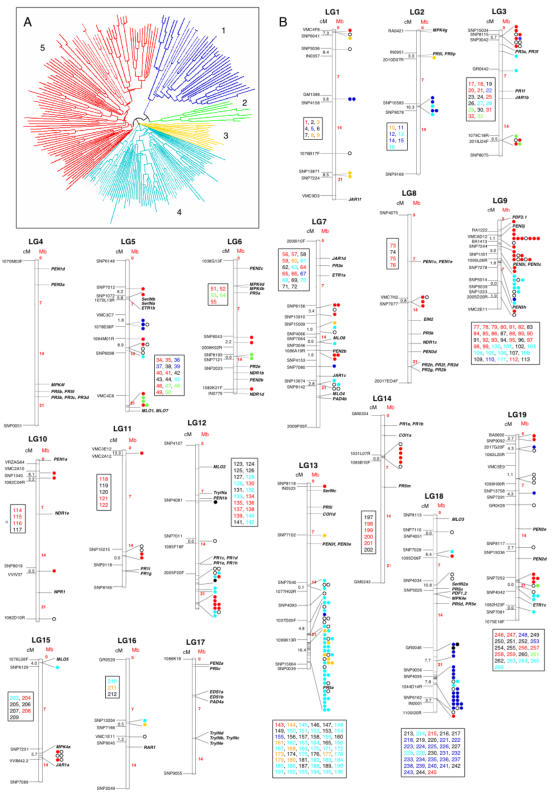
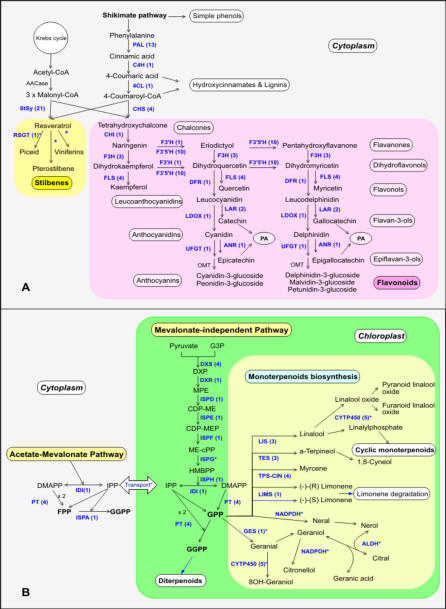
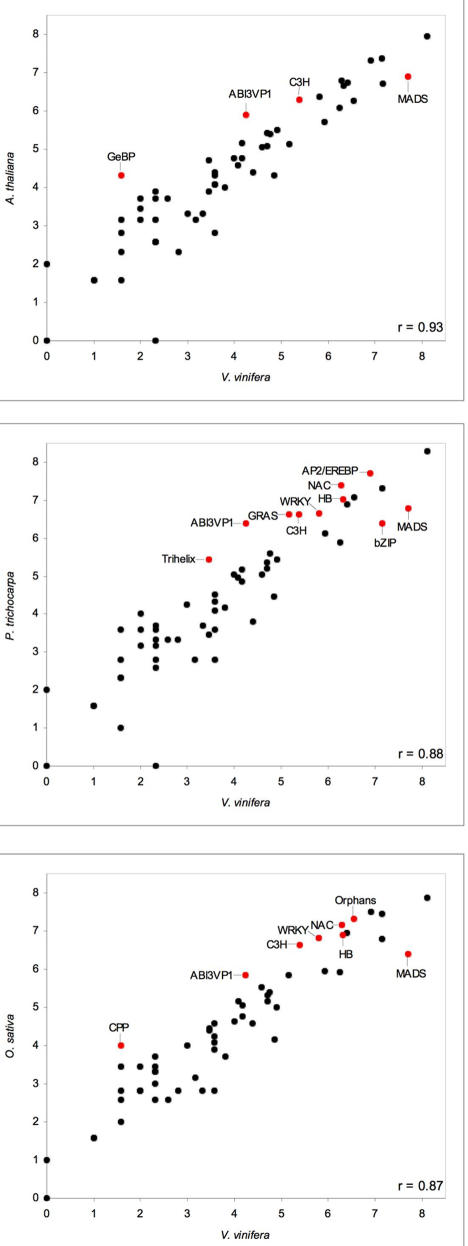
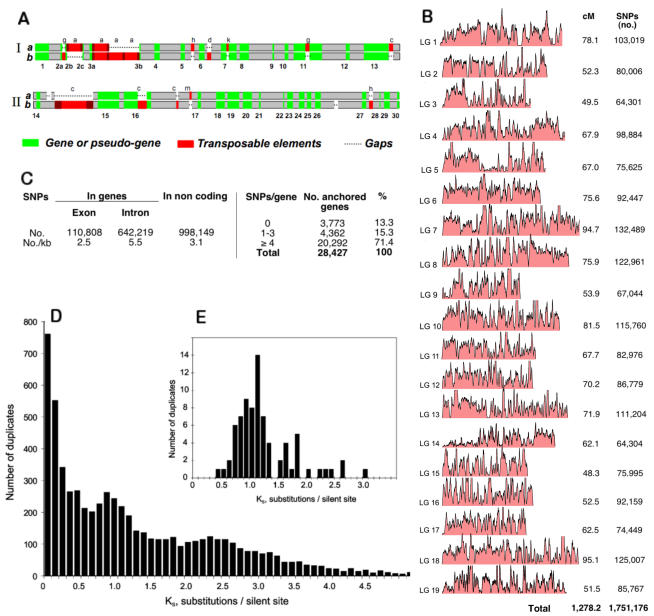
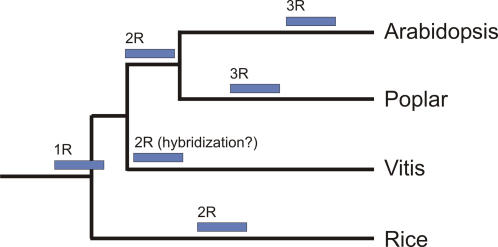
Principal findings: We estimate the genome size of V. vinifera to be 504.6 Mb. Genomic sequences corresponding to 477.1 Mb were assembled in 2,093 metacontigs and 435.1 Mb were anchored to the 19 linkage groups (LGs). The number of predicted genes is 29,585, of which 96.1% were assigned to LGs. This assembly of the grape genome provides candidate genes implicated in traits relevant to grapevine cultivation, such as those influencing wine quality, via secondary metabolites, and those connected with the extreme susceptibility of grape to pathogens. Single nucleotide polymorphism (SNP) distribution was consistent with a diffuse haplotype structure across the genome. Of around 2,000,000 SNPs, 1,751,176 were mapped to chromosomes and one or more of them were identified in 86.7% of anchored genes. The relative age of grape duplicated genes was estimated and this made possible to reveal a relatively recent Vitis-specific large scale duplication event concerning at least 10 chromosomes (duplication not reported before).
Conclusions: Sanger shotgun sequencing and highly efficient sequencing by synthesis (SBS), together with dedicated assembly programs, resolved a complex heterozygous genome. A consensus sequence of the genome and a set of mapped marker loci were generated. Homologous chromosomes of Pinot Noir differ by 11.2% of their DNA (hemizygous DNA plus chromosomal gaps). SNP markers are offered as a tool with the potential of introducing a new era in the molecular breeding of grape.
Conflict of interest statement
Figures

References
-
- McGovern PE. 2003. Ancient Wine: The Search for the Origins of Viniculture Princeton University Press, Princeton.
-
- Panagiotakos DB, Pitsavos C, Polychronopoulos E, Chrysohoou C, Zampelas A, et al. Can a Mediterranean diet moderate the development and clinical progression of coronary heart disease? A systematic review. Med Sci Monit. 2004;10:RA193–198. - PubMed
-
- Burns J, Gardner PT, O'Neil J, Crawford S, Morecroft I, et al. Relationship among Antioxidant Activity, Vasodilation Capacity, and Phenolic Content of Red Wines. J Agric Food Chem. 2000;48:220–230. - PubMed
-
- Levadoux L. Les populations sauvages et cultivées de Vitis vinifera L. Ann Amélior Plantes. 1956;6:59–117.
-
- Olmo HP. Grapes. In: Simmonds NW, editor. Evolution of crop plants. London: 1979.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
