

Conduction velocity is regulated by sodium channel inactivation in unmyelinated axons innervating the rat cranial meninges
- PMID: 18096592
- PMCID: PMC2375633
- DOI: 10.1113/jphysiol.2007.145383
Conduction velocity is regulated by sodium channel inactivation in unmyelinated axons innervating the rat cranial meninges
Abstract
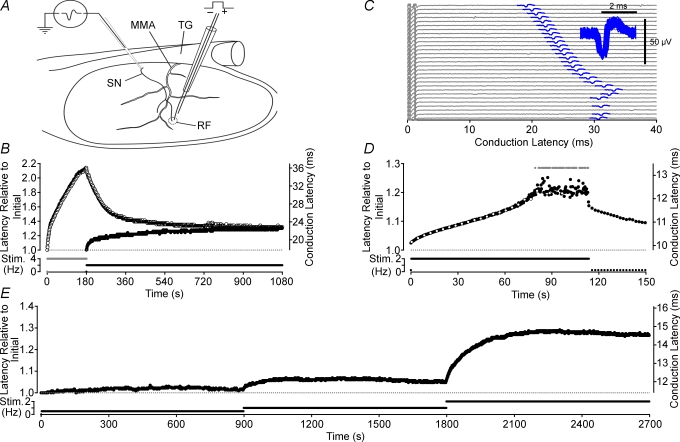
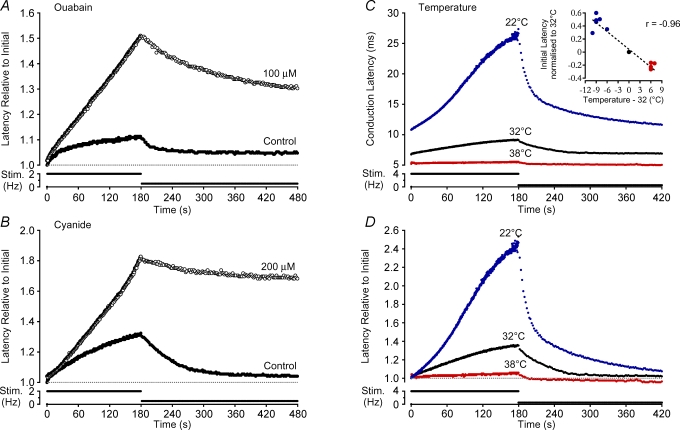
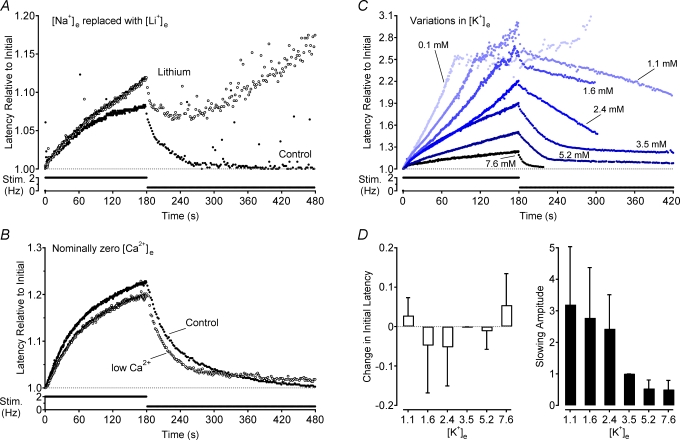
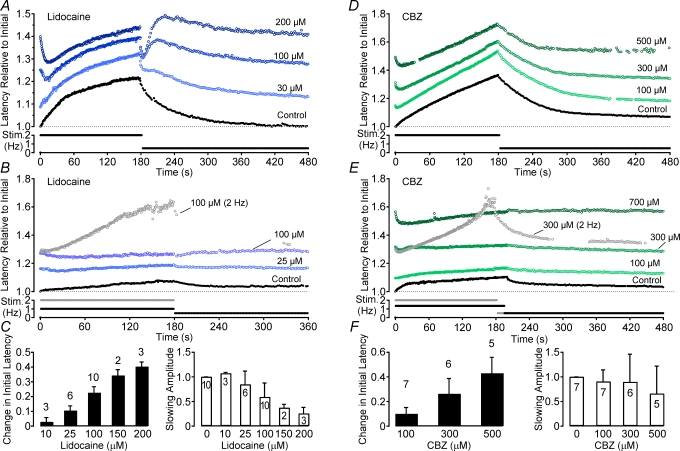
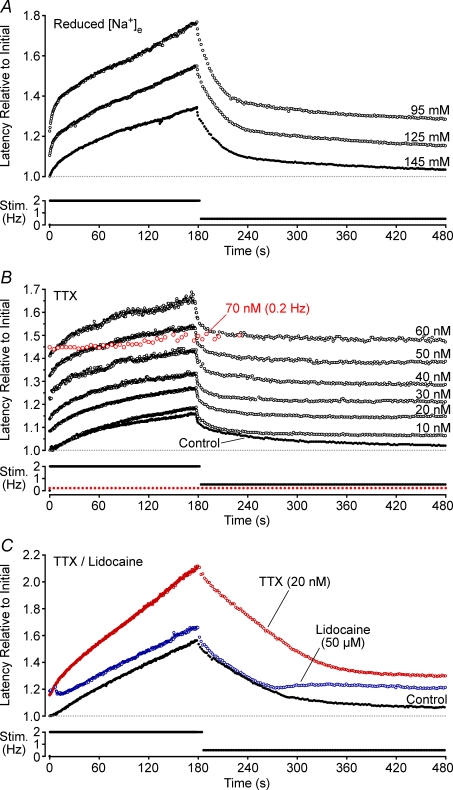
Axonal conduction velocity varies according to the level of preceding impulse activity. In unmyelinated axons this typically results in a slowing of conduction velocity and a parallel increase in threshold. It is currently held that Na(+)-K(+)-ATPase-dependent axonal hyperpolarization is responsible for this slowing but this has long been equivocal. We therefore examined conduction velocity changes during repetitive activation of single unmyelinated axons innervating the rat cranial meninges. In direct contradiction to the currently accepted postulate, Na(+)-K(+)-ATPase blockade actually enhanced activity-induced conduction velocity slowing, while the degree of velocity slowing was curtailed in the presence of lidocaine (10-300 microm) and carbamazepine (30-500 microm) but not tetrodotoxin (TTX, 10-80 nm). This suggests that a change in the number of available sodium channels is the most prominent factor responsible for activity-induced changes in conduction velocity in unmyelinated axons. At moderate stimulus frequencies, axonal conduction velocity is determined by an interaction between residual sodium channel inactivation following each impulse and the retrieval of channels from inactivation by a concomitant Na(+)-K(+)-ATPase-mediated hyperpolarization. Since the process is primarily dependent upon sodium channel availability, tracking conduction velocity provides a means of accessing relative changes in the excitability of nociceptive neurons.
Figures
Comment in
-
Evidence of a physiological role for use-dependent inactivation of NaV1.8 sodium channels.J Physiol. 2008 Feb 15;586(4):923. doi: 10.1113/jphysiol.2008.150821. J Physiol. 2008. PMID: 18287386 Free PMC article. No abstract available.
References
-
- Akopian AN, Sivilotti L, Wood JN. A tetrodotoxin-resistant voltage-gated sodium channel expressed by sensory neurons. Nature. 1996;379:257–262. - PubMed
-
- Andres KH, von During M, Muszynski K, Schmidt RF. Nerve fibres and their terminals of the dura mater encephali of the rat. Anat Embryol (Berl) 1987;175:289–301. - PubMed
-
- Beck PW, Handwerker HO, Zimmermann M. Nervous outflow from the cat/s foot during noxious radiant heat stimulation. Brain Res. 1974;67:373–386. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources