

Phosphorylation of vascular endothelial cadherin controls lymphocyte emigration
- PMID: 18096689
- PMCID: PMC3810954
- DOI: 10.1242/jcs.022681
Phosphorylation of vascular endothelial cadherin controls lymphocyte emigration
Abstract
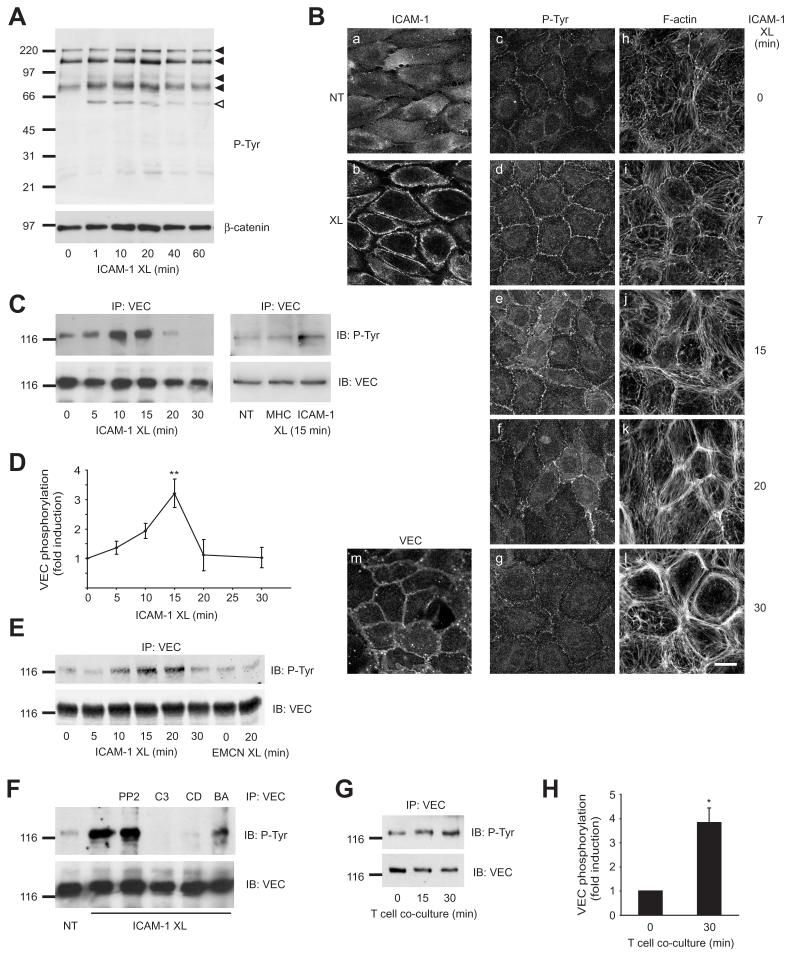
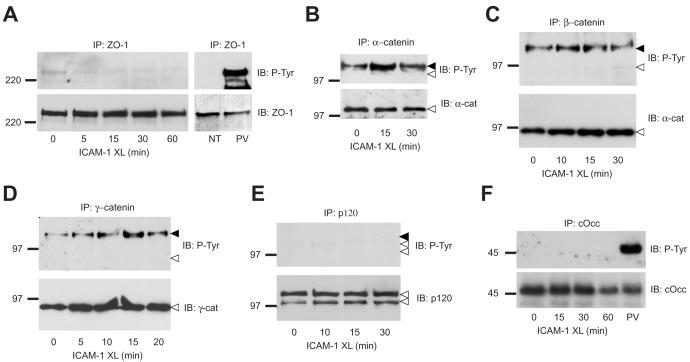
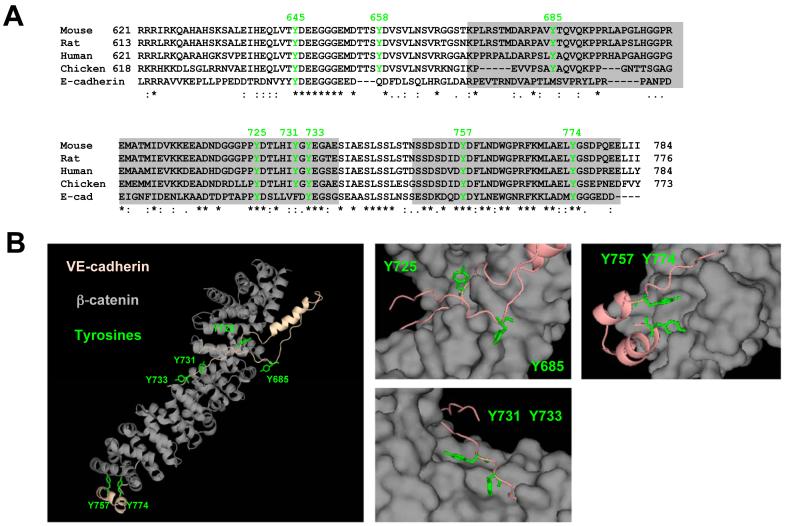
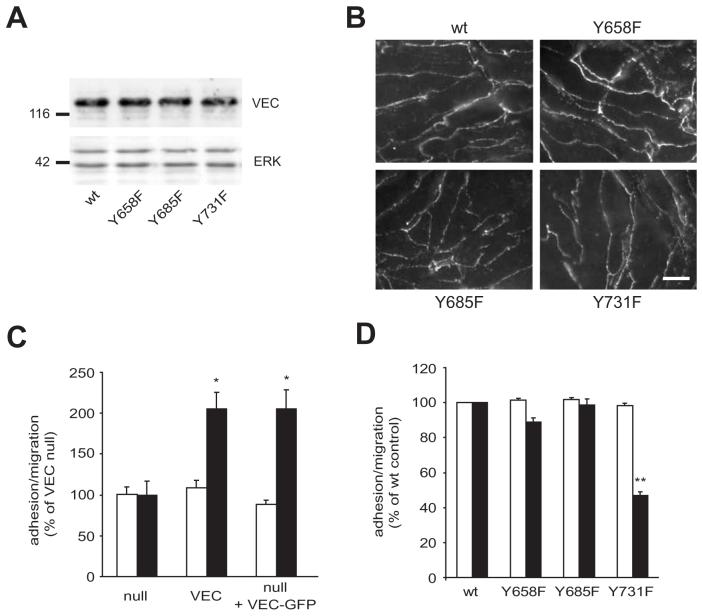
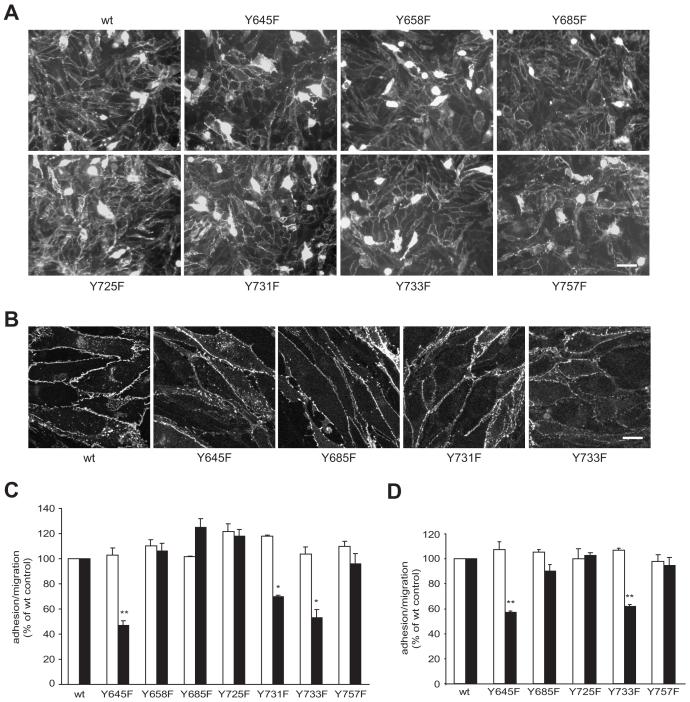
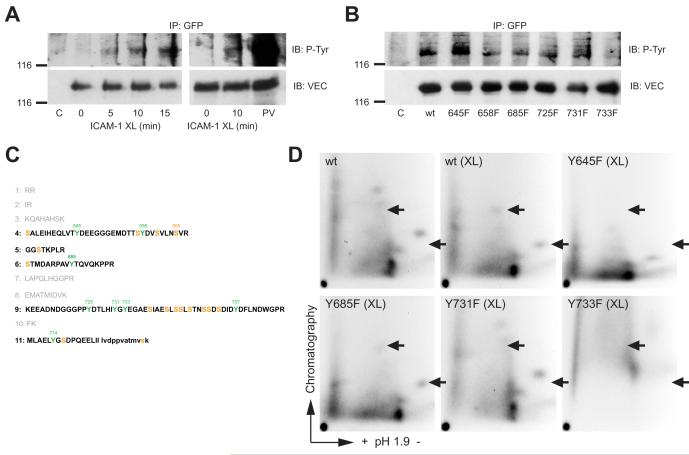
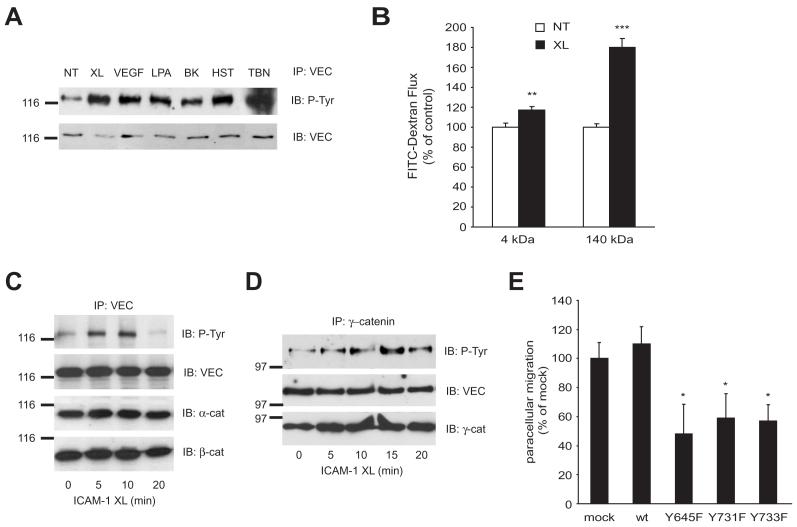
Lymphocytes emigrate from the circulation to target tissues through the microvascular endothelial cell (EC) barrier. During paracellular transmigration cell-cell junctions have been proposed to disengage and provide homophilic and heterophilic interaction surfaces in a zip-like process. However, it is not known whether ECs modulate junction proteins during this process. Here we show that tyrosine phosphorylation of adherens junction vascular endothelial cadherin (VEC) is required for successful transendothelial lymphocyte migration. We found that adhesion of lymphocytes or activation of the endothelial intercellular adhesion molecule 1 (ICAM1) led to tyrosine phosphorylation of VEC. Substitution of tyrosine for phenylalanine in VEC at positions 645, 731 or 733 produced ECs that were significantly less permissive to lymphocyte migration. We also found that these same tyrosine residues are involved in ICAM1-dependent changes of VEC phosphorylation. ICAM1 activation enhanced transendothelial permeability, suggesting the occurrence of junction disassembly. In agreement, the expression of VEC mutated at Y645F, Y731F or Y733F predominantly affected lymphocyte transmigration in paracellular areas. Taken together, these results demonstrate that phosphorylation of adherens junctions constitutes a molecular endpoint of lymphocyte-induced vascular EC signaling and may be exploited as a new target of anti-inflammatory therapies.
Figures
References
-
- Adamson P, Etienne S, Couraud PO, Calder V, Greenwood J. Lymphocyte migration through brain endothelial cell monolayers involves signaling through endothelial ICAM-1 via a rho-dependent pathway. J. Immunol. 1999;162:2964–2973. - PubMed
-
- Allingham MJ, van Buul JD, Burridge K. ICAM-1-Mediated, Src- and Pyk2-Dependent Vascular Endothelial Cadherin Tyrosine Phosphorylation Is Required for Leukocyte Transendothelial Migration. J. Immunol. 2007;179:4053–4064. - PubMed
-
- Andriopoulou P, Navarro P, Zanetti A, Lampugnani MG, Dejana E. Histamine induces tyrosine phosphorylation of endothelial cell-to-cell adherens junctions. Arterioscler. Thromb. Vasc. Biol. 1999;19:2286–2297. - PubMed
-
- Bazzoni G, Dejana E. Endothelial cell-to-cell junctions: molecular organization and role in vascular homeostasis. Physiol Rev. 2004;84:869–901. - PubMed
-
- Boyle WJ, van der GP, Hunter T. Phosphopeptide mapping and phosphoamino acid analysis by two-dimensional separation on thin-layer cellulose plates. Methods Enzymol. 1991;201:110–149. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
