

Controlled exchange of chromosomal arms reveals principles driving telomere interactions in yeast
- PMID: 18096749
- PMCID: PMC2203624
- DOI: 10.1101/gr.6687808
Controlled exchange of chromosomal arms reveals principles driving telomere interactions in yeast
Abstract
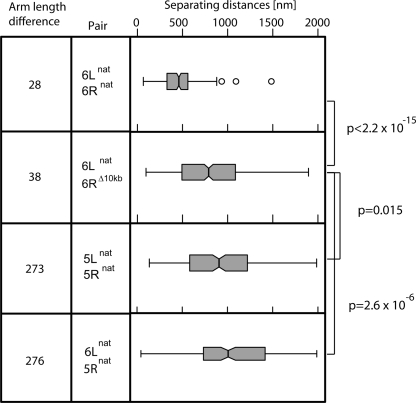
The 32 telomeres in the budding yeast genome cluster in three to seven perinuclear foci. Although individual telomeres and telomeric foci are in constant motion, preferential juxtaposition of some telomeres has been scored. To examine the principles that guide such long-range interactions, we differentially tagged pairs of chromosome ends and developed an automated three-dimensional measuring tool that determines distances between two telomeres. In yeast, all chromosomal ends terminate in TG(1-3) and middle repetitive elements, yet subgroups of telomeres also share extensive homology in subtelomeric coding domains. We find that up to 21 kb of >90% sequence identity does not promote telomere pairing in interphase cells. To test whether unique sequence elements, arm length, or chromosome territories influence juxtaposition, we reciprocally swapped terminal domains or entire chromosomal arms from one chromosome to another. We find that the distal 10 kb of Tel6R promotes interaction with Tel6L, yet only when the two telomeres are present on the same chromosome. By manipulating the length and sequence composition of the right arm of chr 5, we confirm that contact between telomeres on opposite chromatid arms of equal length is favored. These results can be explained by the polarized Rabl arrangement of yeast centromeres and telomeres, which promote to telomere pairing by allowing contact between chromosome arms of equal length in anaphase.
Figures






References
-
- Andrulis E.D., Neiman A.M., Zappulla D.C., Sternglanz R., Neiman A.M., Zappulla D.C., Sternglanz R., Zappulla D.C., Sternglanz R., Sternglanz R. Perinuclear localization of chromatin facilitates transcriptional silencing. Nature. 1998;394:592–595. - PubMed
-
- Burgess-Beusse B., Farrell C., Gaszner M., Litt M., Mutskov V., Recillas-Targa F., Simpson M., West A., Felsenfeld G., Farrell C., Gaszner M., Litt M., Mutskov V., Recillas-Targa F., Simpson M., West A., Felsenfeld G., Gaszner M., Litt M., Mutskov V., Recillas-Targa F., Simpson M., West A., Felsenfeld G., Litt M., Mutskov V., Recillas-Targa F., Simpson M., West A., Felsenfeld G., Mutskov V., Recillas-Targa F., Simpson M., West A., Felsenfeld G., Recillas-Targa F., Simpson M., West A., Felsenfeld G., Simpson M., West A., Felsenfeld G., West A., Felsenfeld G., Felsenfeld G. The insulation of genes from external enhancers and silencing chromatin. Proc. Natl. Acad. Sci. 2002;99:16433–16437. (Suppl. 4) - PMC - PubMed
-
- Bystricky K., Heun P., Gehlen L., Langowski J., Gasser S.M., Heun P., Gehlen L., Langowski J., Gasser S.M., Gehlen L., Langowski J., Gasser S.M., Langowski J., Gasser S.M., Gasser S.M. Long-range compaction and flexibility of interphase chromatin in budding yeast analyzed by high-resolution imaging techniques. Proc. Natl. Acad. Sci. 2004;101:16495–16500. - PMC - PubMed
-
- Bystricky K., Laroche T., van Houwe G., Blaszczyk M., Gasser S.M., Laroche T., van Houwe G., Blaszczyk M., Gasser S.M., van Houwe G., Blaszczyk M., Gasser S.M., Blaszczyk M., Gasser S.M., Gasser S.M. Chromosome looping in yeast: Telomere pairing and coordinated movement reflect anchoring efficiency and territorial organization. J. Cell Biol. 2005;168:375–387. - PMC - PubMed
-
- Celniker S.E., Drewell R.A., Drewell R.A. Chromatin looping mediates boundary element promoter interactions. Bioessays. 2007;29:7–10. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous