

Distinctions among GABAA and GABAB responses revealed by calcium channel antagonists, cannabinoids, opioids, and synaptic plasticity in rat hippocampus
- PMID: 18097653
- PMCID: PMC2906116
- DOI: 10.1007/s00213-007-1040-4
Distinctions among GABAA and GABAB responses revealed by calcium channel antagonists, cannabinoids, opioids, and synaptic plasticity in rat hippocampus
Abstract
Rationale: Hippocampal interneurons release gamma-aminobutyric acid (GABA) and produce fast GABA(A)- and slow GABA(B)-inhibitory postsynaptic potentials (IPSPs). The regulation of GABA(B) eIPSPs or the interneurons that produce them are not well understood. In addition, while both micro-opioid receptors (microORs) and cannabinoid CB1R receptors (CB1Rs) are present on hippocampal interneurons, it is not clear how these two systems interact.
Objectives: This study tests the hypotheses that: (1) all interneurons can initiate both GABA(A) and GABA(B) inhibitory postsynaptic potentials; (2) GABA(B) responses are insensitive to mGluR-triggered, endocannabinoid (eCB)-mediated inhibitory long-term depression (iLTD); (3) GABA(B) responses are produced by interneurons that express microOR; and (4) CB1R-dependent and microOR-dependent response interact.
Materials and methods: Pharmacological and electrophysiological approaches were used in acute rat hippocampal slices. High resistance microelectrode recordings were made from pyramidal cells, while interneurons were stimulated extracellularly.
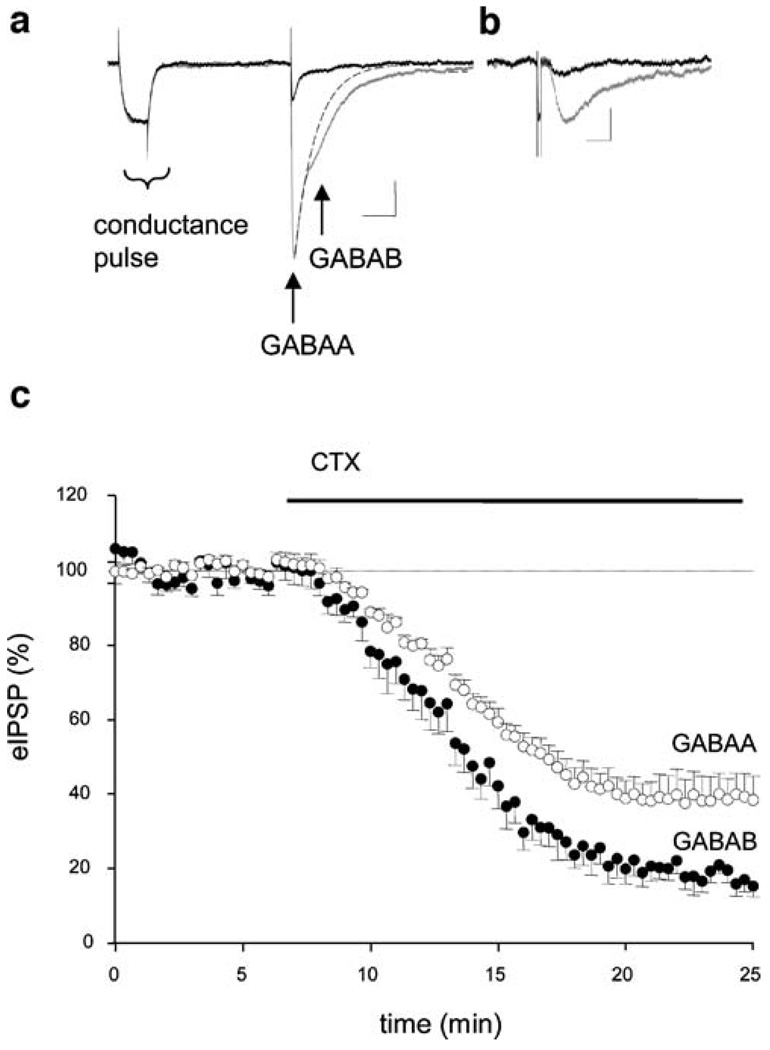
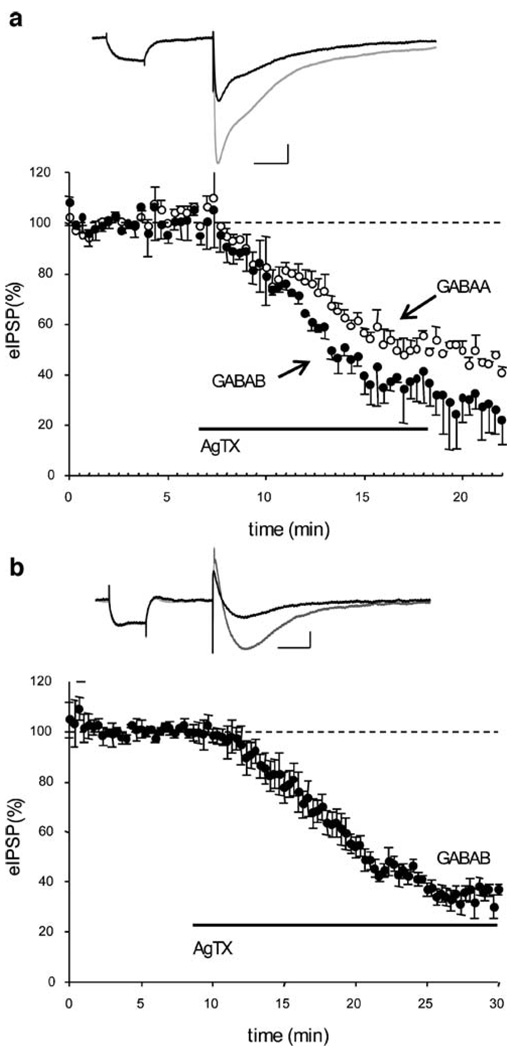
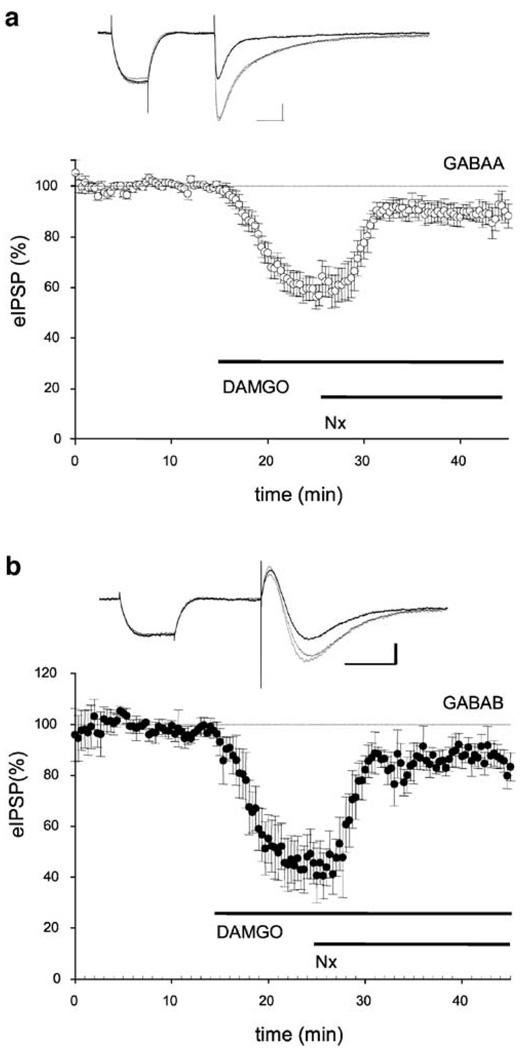
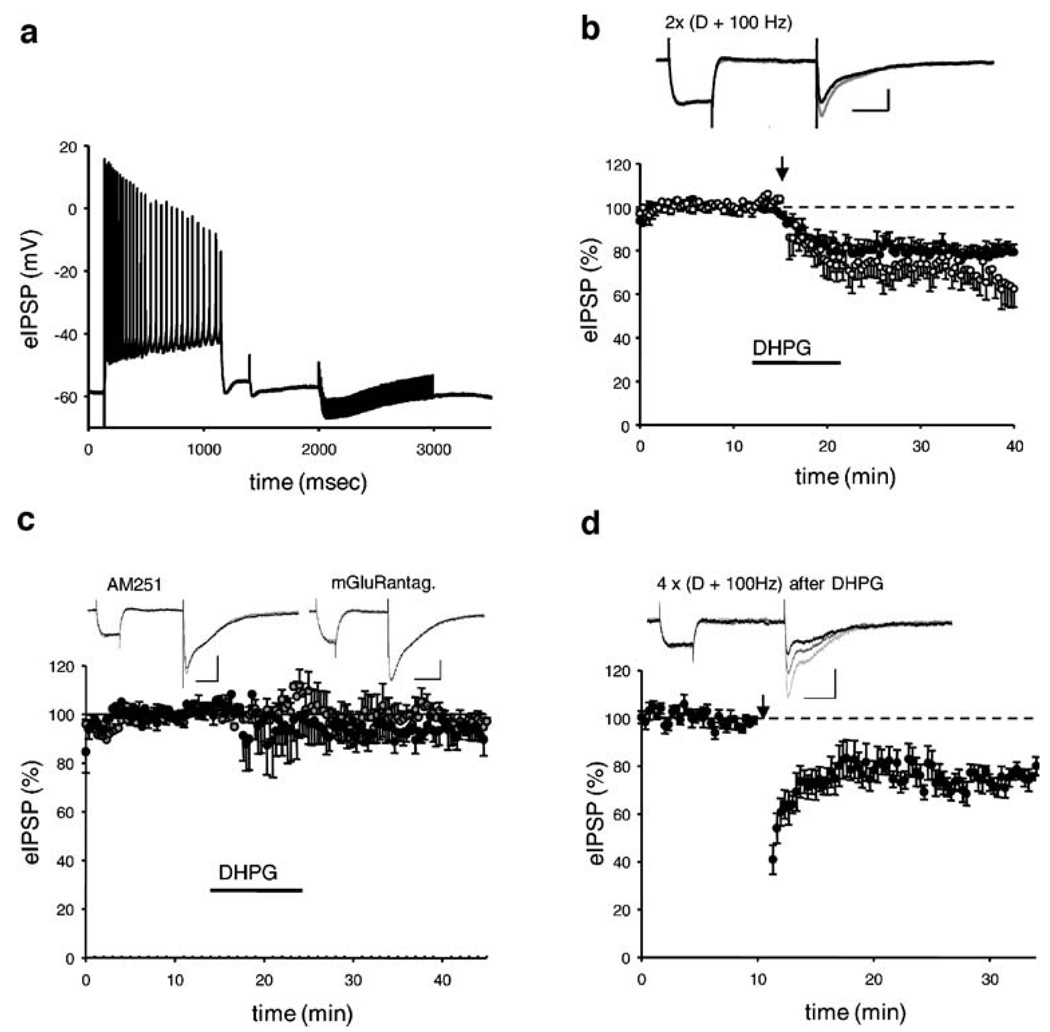
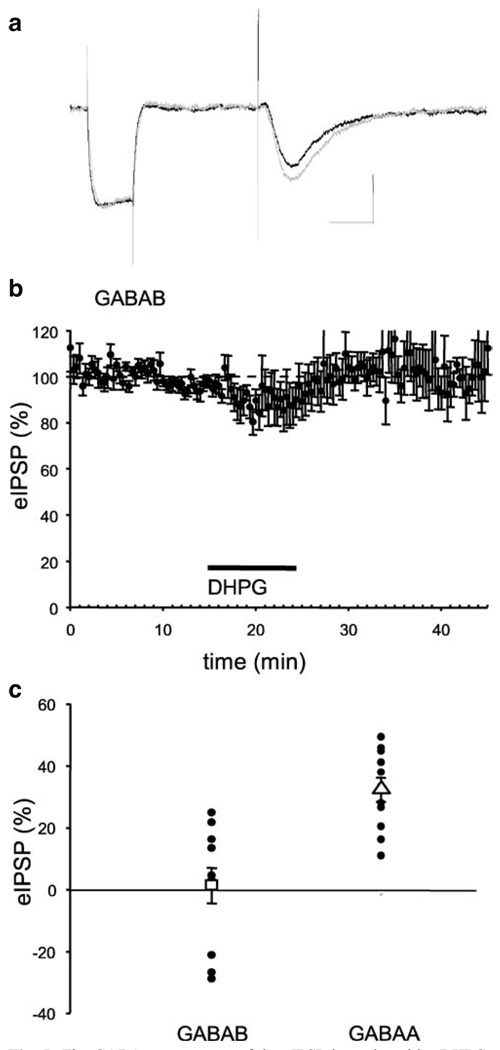
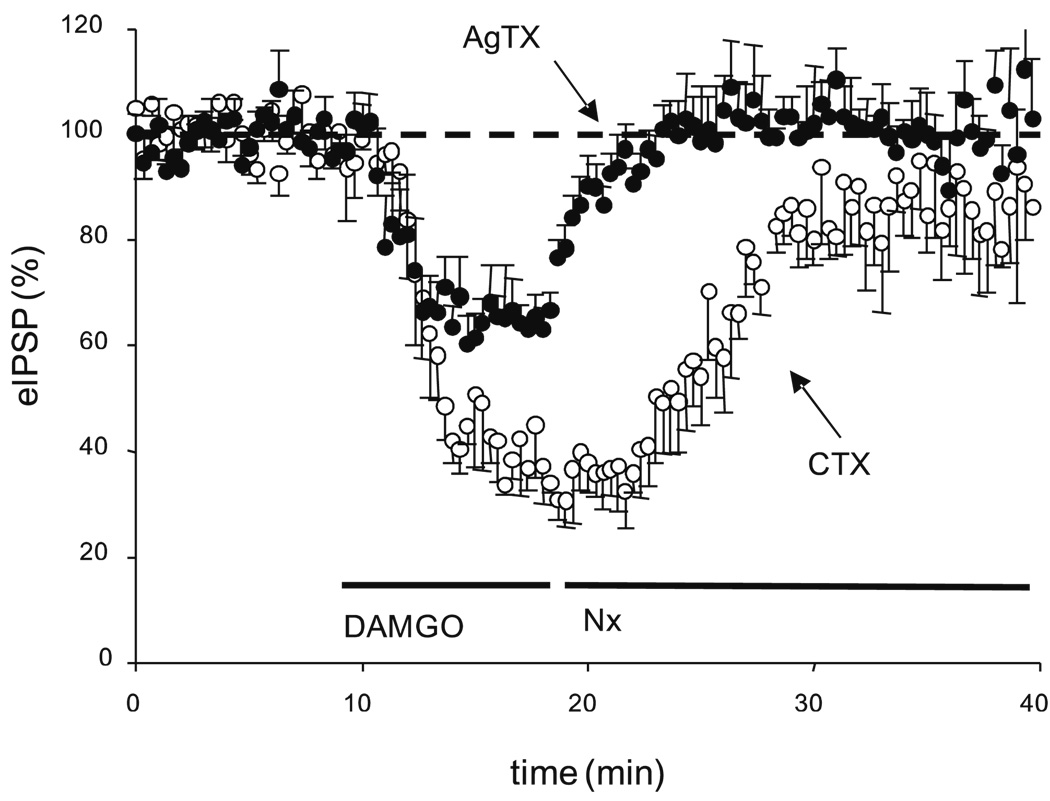
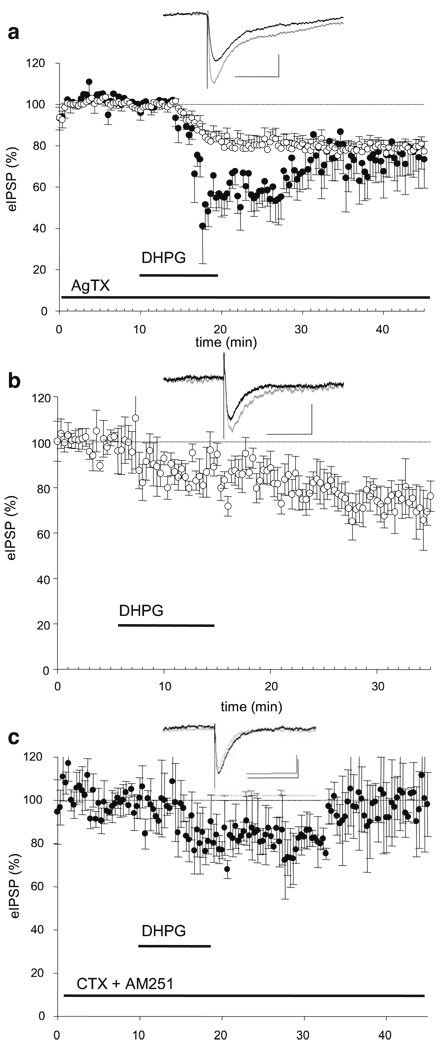
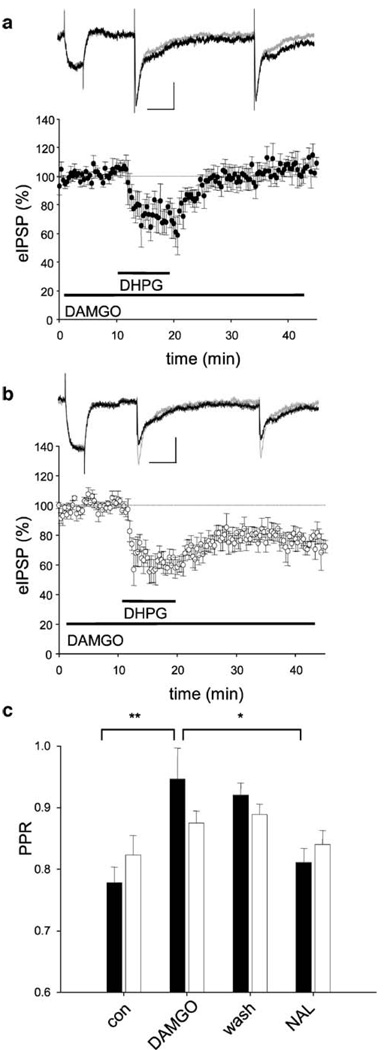
Results: GABA(B) responses were found to be produced by interneurons that release GABA via either presynaptic N-type or P/Q-type calcium channels but that they are insensitive to suppression by eCBs or eCB-mediated iLTD. GABA(B) IPSPs were sensitive to suppression by a microOR agonist, suggesting a major source of GABA(B) responses is the microOR-expressing interneuron population. A small eCB-iLTD (10% eIPSP reduction) persisted in conotoxin. eCB-iLTD was blocked by a microOR agonist in 6/13 slices.
Conclusions: GABA(B) responses cannot be produced by all interneurons. CB1R or microOR agonists will differentially alter the balance of activity in hippocampal circuits. CB1R- and microOR-mediated responses can interact.
Figures
References
-
- Alger BE. Retrograde signaling in the regulation of synaptic transmission: focus on endocannabinoids. Prog Neurobiol. 2002;68:247–286. - PubMed
-
- Andrade R, Malenka RC, Nicoll RA. A G protein couples serotonin and GABAB receptors to the same channels in hippocampus. Science. 1986;234:1261–1265. - PubMed
-
- Bormann J. Electrophysiology of GABAA and GABAB receptor subtypes. Trends Neurosci. 1988;11:112–116. - PubMed
-
- Bowery N. GABAB receptors and their significance in mammalian pharmacology. Trends Pharmacol Sci. 1989;10:401–407. - PubMed
-
- Buzsaki G. Theta oscillations in the hippocampus. Neuron. 2002;33:325–340. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
