

Cell type-specific cleavage of nucleocapsid protein by effector caspases during SARS coronavirus infection
- PMID: 18155731
- PMCID: PMC7094231
- DOI: 10.1016/j.jmb.2007.11.081
Cell type-specific cleavage of nucleocapsid protein by effector caspases during SARS coronavirus infection
Abstract
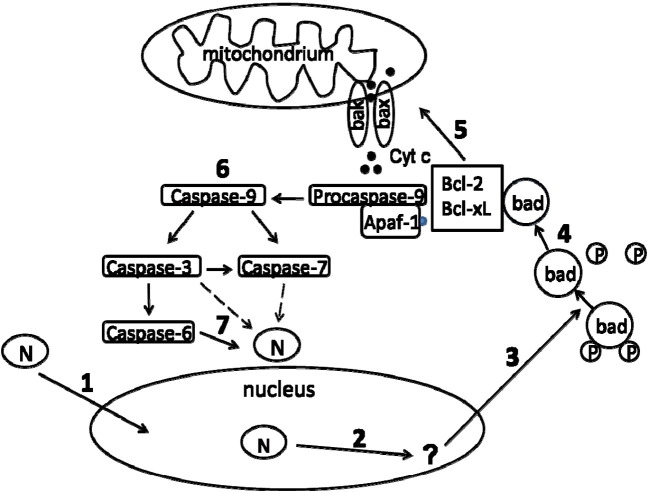
The epidemic outbreak of severe acute respiratory syndrome (SARS) in 2003 was caused by a novel coronavirus (CoV), designated SARS-CoV. The RNA genome of SARS-CoV is complexed by the nucleocapsid protein (N) to form a helical nucleocapsid. Besides this primary function, N seems to be involved in apoptotic scenarios. We show that upon infection of Vero E6 cells with SARS-CoV, which elicits a pronounced cytopathic effect and a high viral titer, N is cleaved by caspases. In contrast, in SARS-CoV-infected Caco-2 cells, which show a moderate cytopathic effect and a low viral titer, this processing of N was not observed. To further verify these observations, we transiently expressed N in different cell lines. Caco-2 and N2a cells served as models for persistent SARS-CoV infection, whereas Vero E6 and A549 cells did as prototype cell lines lytically infected by SARS-CoV. The experiments revealed that N induces the intrinsic apoptotic pathway, resulting in processing of N at residues 400 and 403 by caspase-6 and/or caspase-3. Of note, caspase activation is highly cell type specific in SARS-CoV-infected as well as transiently transfected cells. In Caco-2 and N2a cells, almost no N-processing was detectable. In Vero E6 and A549 cells, a high proportion of N was cleaved by caspases. Moreover, we examined the subcellular localization of SARS-CoV N in these cell lines. In transfected Vero E6 and A549 cells, SARS-CoV N was localized both in the cytoplasm and nucleus, whereas in Caco-2 and N2a cells, nearly no nuclear localization was observed. In addition, our studies indicate that the nuclear localization of N is essential for its caspase-6-mediated cleavage. These data suggest a correlation among the replication cycle of SARS-CoV, subcellular localization of N, induction of apoptosis, and the subsequent activation of caspases leading to cleavage of N.
Figures





References
-
- Drosten C., Gunther S., Preiser W., van der W.S., Brodt H.R., Becker S. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N. Engl. J. Med. 2003;348:1967–1976. - PubMed
-
- Ksiazek T.G., Erdman D., Goldsmith C.S., Zaki S.R., Peret T., Emery S. A novel coronavirus associated with severe acute respiratory syndrome. N. Engl. J. Med. 2003;348:1953–1966. - PubMed
-
- Marra M.A., Jones S.J., Astell C.R., Holt R.A., Brooks-Wilson A., Butterfield Y.S. The genome sequence of the SARS-associated coronavirus. Science. 2003;300:1399–1404. - PubMed
-
- Rota P.A., Oberste M.S., Monroe S.S., Nix W.A., Campagnoli R., Icenogle J.P. Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science. 2003;300:1394–1399. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
