

Role of heme in the antifungal activity of the azaoxoaporphine alkaloid sampangine
- PMID: 18156292
- PMCID: PMC2238161
- DOI: 10.1128/EC.00323-07
Role of heme in the antifungal activity of the azaoxoaporphine alkaloid sampangine
Abstract
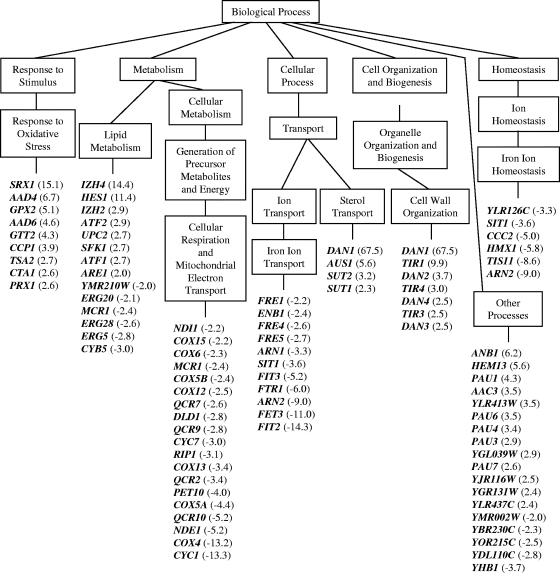
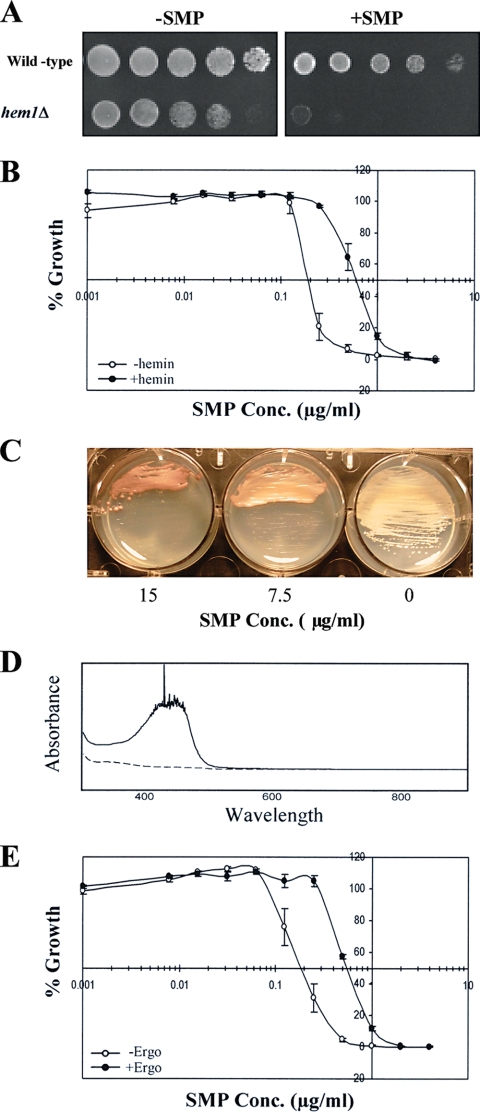
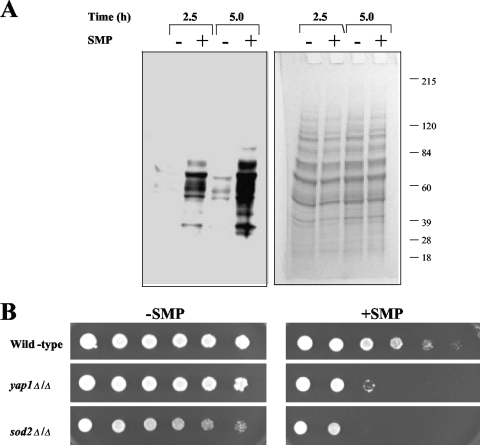
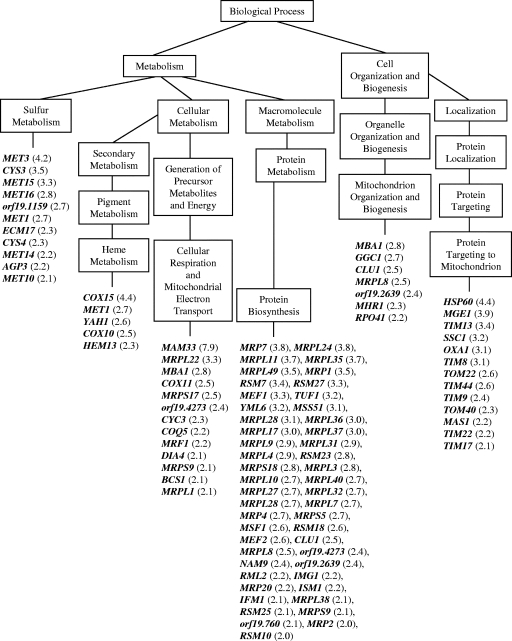
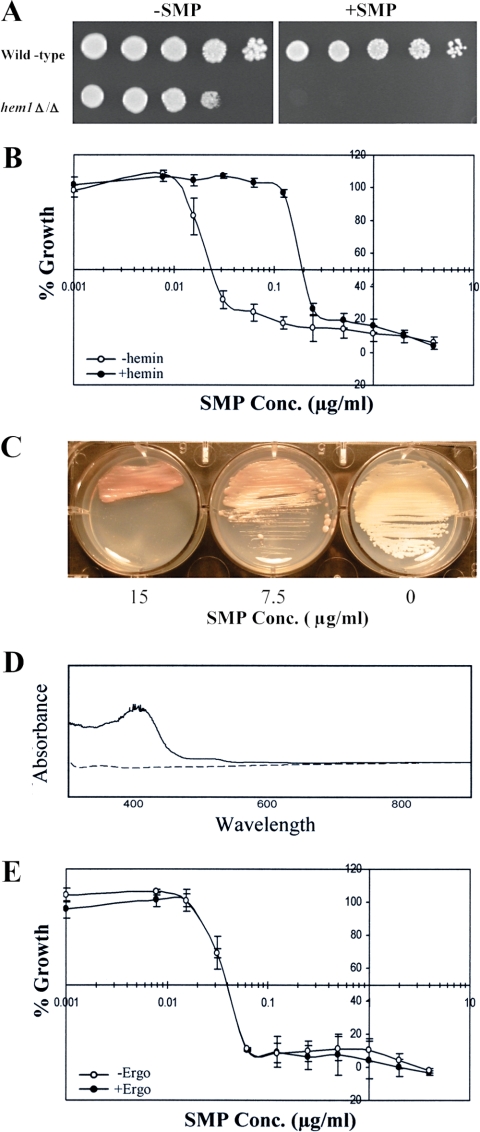
Sampangine, a plant-derived alkaloid found in the Annonaceae family, exhibits strong inhibitory activity against the opportunistic fungal pathogens Candida albicans, Cryptococcus neoformans, and Aspergillus fumigatus. In the present study, transcriptional profiling experiments coupled with analyses of mutants were performed in an effort to elucidate its mechanism of action. Using Saccharomyces cerevisiae as a model organism, we show that sampangine produces a transcriptional response indicative of hypoxia, altering the expression of genes known to respond to low-oxygen conditions. Several additional lines of evidence obtained suggest that these responses could involve effects on heme. First, the hem1Delta mutant lacking the first enzyme in the heme biosynthetic pathway showed increased sensitivity to sampangine, and exogenously supplied hemin partially rescued the inhibitory activity of sampangine in wild-type cells. In addition, heterozygous mutants with deletions in genes involved in five out of eight steps in the heme biosynthetic pathway showed increased susceptibility to sampangine. Furthermore, spectral analyses of pyridine extracts indicated significant accumulation of free porphyrins in sampangine-treated cells. Transcriptional profiling experiments were also performed with C. albicans to investigate the response of a pathogenic fungal species to sampangine. Taking into account the known differences in the physiological responses of C. albicans and S. cerevisiae to low oxygen, significant correlations were observed between the two transcription profiles, suggestive of heme-related defects. Our results indicate that the antifungal activity of the plant alkaloid sampangine is due, at least in part, to perturbations in the biosynthesis or metabolism of heme.
Figures


References
-
- Atamna, H., P. B. Walter, and B. N. Ames. 2002. The role of heme and iron-sulfur clusters in mitochondrial biogenesis, maintenance, and decay with age. Arch. Biochem. Biophys. 397345-353. - PubMed
-
- Bard, M., A. M. Sturm, C. A. Pierson, S. Brown, K. M. Rogers, S. Nabinger, J. Eckstein, R. Barbuch, N. D. Lees, S. A. Howell, and K. C. Hazen. 2005. Sterol uptake in Candida glabrata: rescue of sterol auxotrophic strains. Diagn. Microbiol. Infect. Dis. 52285-293. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
