

Salicylic acid and systemic acquired resistance play a role in attenuating crown gall disease caused by Agrobacterium tumefaciens
- PMID: 18156296
- PMCID: PMC2245820
- DOI: 10.1104/pp.107.111302
Salicylic acid and systemic acquired resistance play a role in attenuating crown gall disease caused by Agrobacterium tumefaciens
Abstract
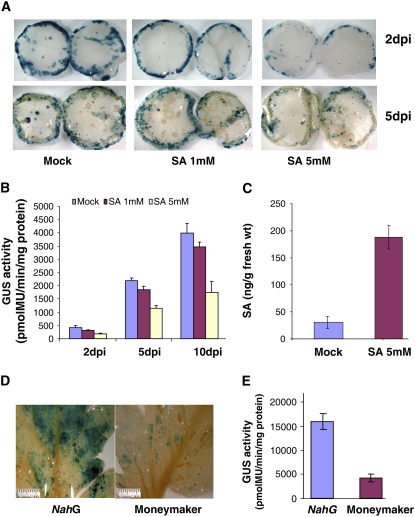
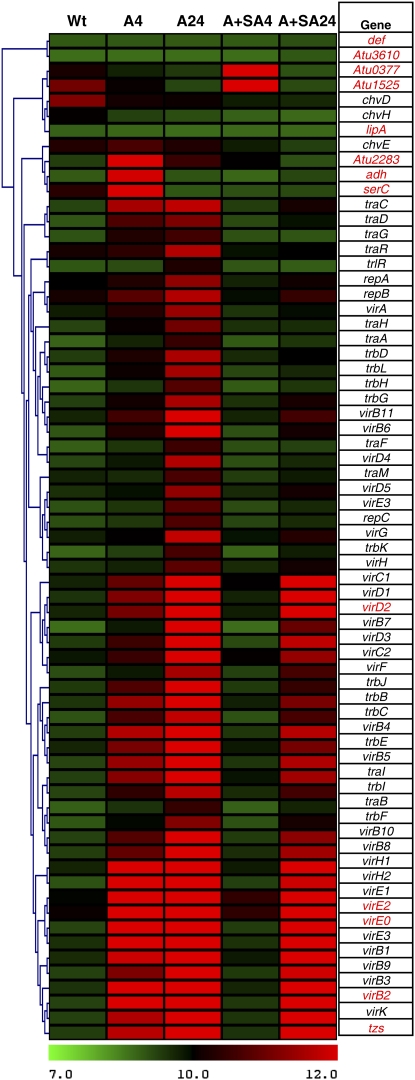
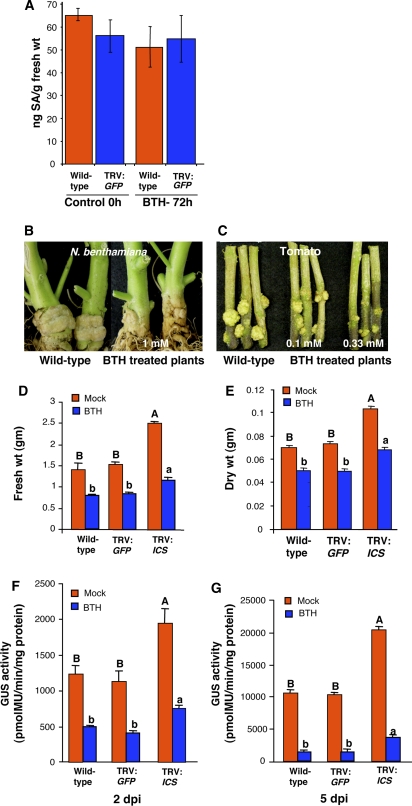
We investigated the effects of salicylic acid (SA) and systemic acquired resistance (SAR) on crown gall disease caused by Agrobacterium tumefaciens. Nicotiana benthamiana plants treated with SA showed decreased susceptibility to Agrobacterium infection. Exogenous application of SA to Agrobacterium cultures decreased its growth, virulence, and attachment to plant cells. Using Agrobacterium whole-genome microarrays, we characterized the direct effects of SA on bacterial gene expression and showed that SA inhibits induction of virulence (vir) genes and the repABC operon, and differentially regulates the expression of many other sets of genes. Using virus-induced gene silencing, we further demonstrate that plant genes involved in SA biosynthesis and signaling are important determinants for Agrobacterium infectivity on plants. Silencing of ICS (isochorismate synthase), NPR1 (nonexpresser of pathogenesis-related gene 1), and SABP2 (SA-binding protein 2) in N. benthamiana enhanced Agrobacterium infection. Moreover, plants treated with benzo-(1,2,3)-thiadiazole-7-carbothioic acid, a potent inducer of SAR, showed reduced disease symptoms. Our data suggest that SA and SAR both play a major role in retarding Agrobacterium infectivity.
Figures


References
-
- Achuo EA, Audenaert K, Meziane H, Hofte M (2004) The salicylic acid-dependent defence pathway is effective against different pathogens in tomato and tobacco. Plant Pathol 53 65–72 - PubMed
-
- Agrios GN (1997) Plant Pathology, Ed 4. Academic Press, San Diego
-
- Anand A, Vaghchhipawala Z, Ryu CM, Kang L, Wang K, del-Pozo O, Martin GB, Mysore KS (2007. b) Identification of plant genes involved in Agrobacterium-mediated transformation by using virus-induced gene silencing as a functional genomics tool. Mol Plant Microbe Interact 20 41–52 - PubMed
-
- Bolstad BM, Irizarry RA, Astrand M, Speed TP (2003) A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 19 185–193 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
