

Diverse roles of actin in C. elegans early embryogenesis
- PMID: 18157918
- PMCID: PMC2323969
- DOI: 10.1186/1471-213X-7-142
Diverse roles of actin in C. elegans early embryogenesis
Abstract
Background: The actin cytoskeleton plays critical roles in early development in Caenorhabditis elegans. To further understand the complex roles of actin in early embryogenesis we use RNAi and in vivo imaging of filamentous actin (F-actin) dynamics.
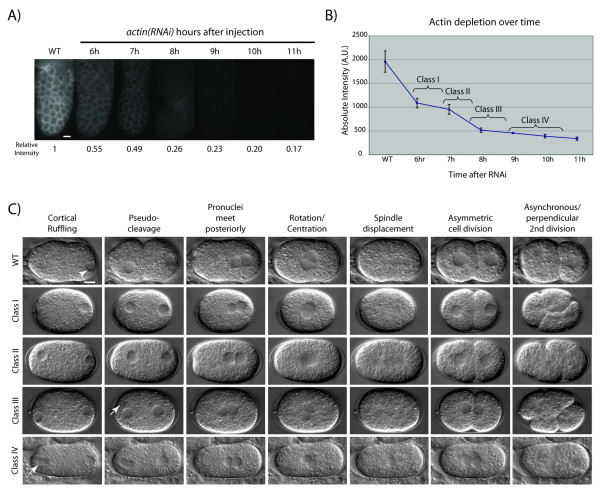
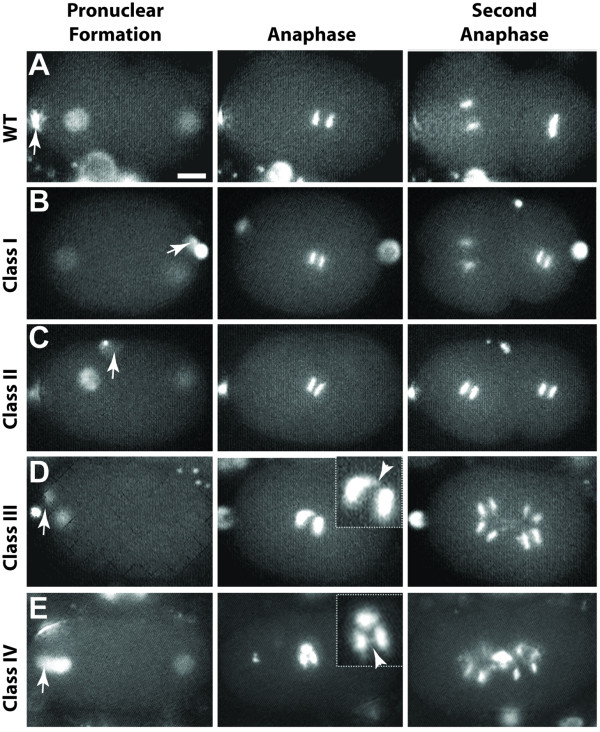
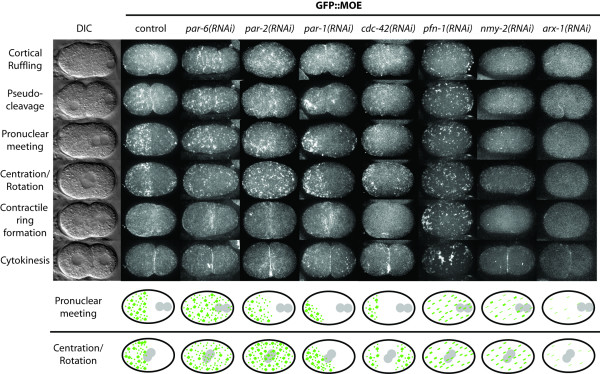
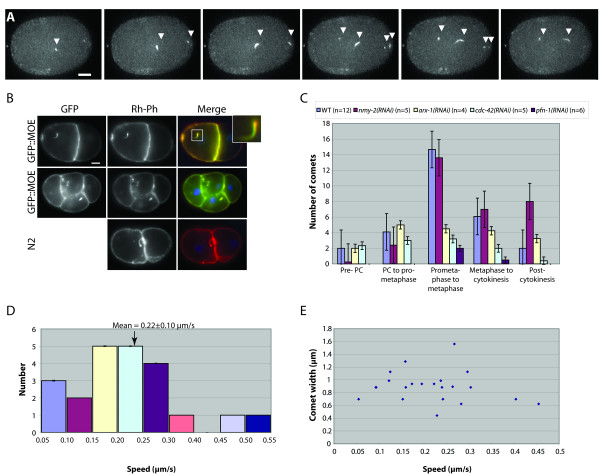
Results: Using RNAi, we found processes that are differentially sensitive to levels of actin during early embryogenesis. Mild actin depletion shows defects in cortical ruffling, pseudocleavage, and establishment of polarity, while more severe depletion shows defects in polar body extrusion, cytokinesis, chromosome segregation, and eventually, egg production. These defects indicate that actin is required for proper oocyte development, fertilization, and a wide range of important events during early embryogenesis, including proper chromosome segregation. In vivo visualization of the cortical actin cytoskeleton shows dynamics that parallel but are distinct from the previously described myosin dynamics. Two distinct types of actin organization are observed at the cortex. During asymmetric polarization to the anterior, or the establishment phase (Phase I), actin forms a meshwork of microfilaments and focal accumulations throughout the cortex, while during the anterior maintenance phase (Phase II) it undergoes a morphological transition to asymmetrically localized puncta. The proper asymmetric redistribution is dependent on the PAR proteins, while both asymmetric redistribution and morphological transitions are dependent upon PFN-1 and NMY-2. Just before cytokinesis, actin disappears from most of the cortex and is only found around the presumptive cytokinetic furrow. Finally, we describe dynamic actin-enriched comets in the early embryo.
Conclusion: During early C. elegans embryogenesis actin plays more roles and its organization is more dynamic than previously described. Morphological transitions of F-actin, from meshwork to puncta, as well as asymmetric redistribution, are regulated by the PAR proteins. Results from this study indicate new insights into the cellular and developmental roles of the actin cytoskeleton.
Figures
References
-
- Alberts B. Molecular biology of the cell. 4th. New York , Garland Science; 2002. p. xxxiv, 1463, [86] p..
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
