

A novel insertion pathway of mitochondrial outer membrane proteins with multiple transmembrane segments
- PMID: 18158327
- PMCID: PMC2373507
- DOI: 10.1083/jcb.200702143
A novel insertion pathway of mitochondrial outer membrane proteins with multiple transmembrane segments
Abstract
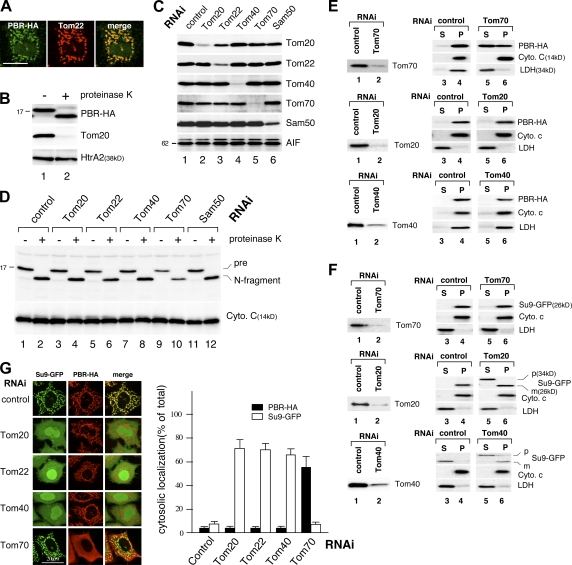
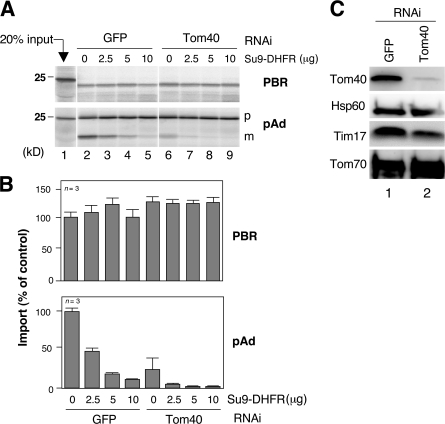
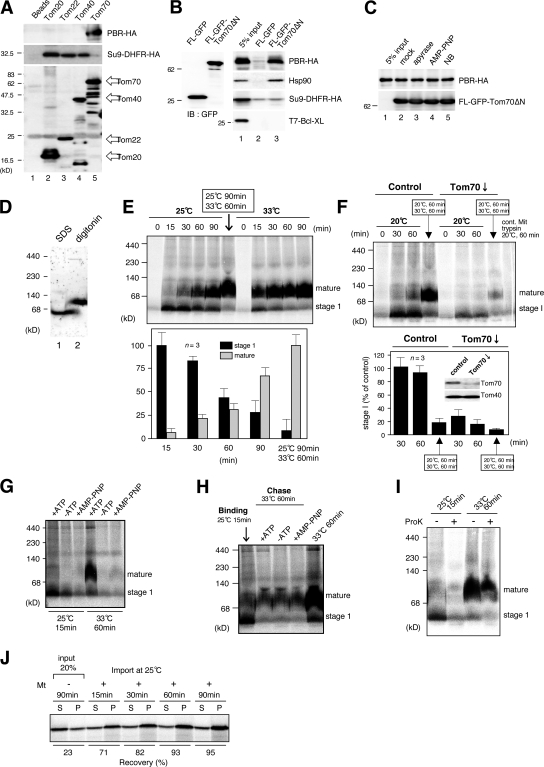
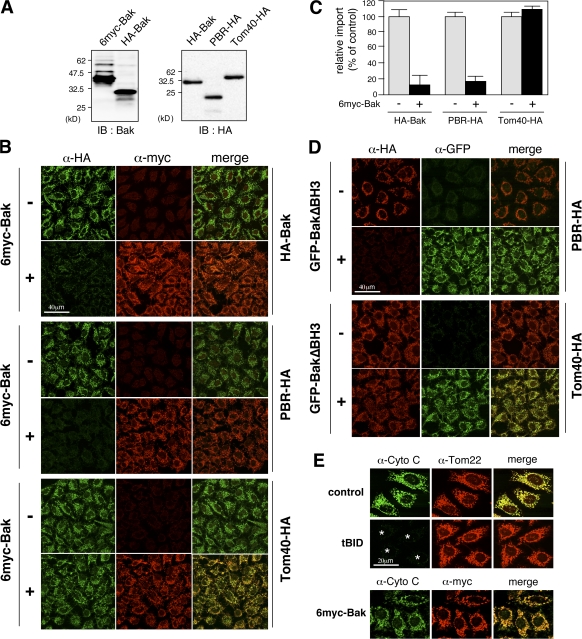
The central channel Tom40 of the preprotein translocase of outer membrane (TOM) complex is thought to be responsible for the import of virtually all preproteins synthesized outside the mitochondria. In this study, we analyze the topogenesis of the peripheral benzodiazepine receptor (PBR), which integrates into the mitochondrial outer membrane (MOM) through five hydrophobic transmembrane segments (TMSs) and functions in cholesterol import into the inner membrane. Analyses of in vitro and in vivo import into TOM component-depleted mitochondria reveal that PBR import (1) depends on the import receptor Tom70 but requires neither the Tom20 and Tom22 import receptors nor the import channel Tom40, (2) shares the post-Tom70 pathway with the C-tail-anchored proteins, and (3) requires factors of the mitochondrial intermembrane space. Furthermore, membrane integration of mitofusins and mitochondrial ubiquitin ligase, the MOM proteins with two and four TMSs, respectively, proceeds through the same initial pathway. These findings reveal a previously unidentified pathway of the membrane integration of MOM proteins with multiple TMSs.
Figures
References
-
- Ahting, U., T. Weizenegger, W. Neupert, and D. Rapaport. 2005. Signal-anchor proteins follow a unique insertion pathway into the outer membrane of mitochondria. J. Biol. Chem. 280:48–53. - PubMed
-
- Bohnert, M., N. Pfanner, and M. van der Laan. 2007. A dynamic machinery for import of mitochondrial precursor proteins. FEBS Lett. 581:2802–2810. - PubMed
-
- Eura, Y., N. Ishihara, S. Yokota, and K. Mihara. 2003. Two mitofusin proteins, mammalian homologues of FZO, with distinct functions are both required for mitochondrial fusion. J. Biochem. (Tokyo). 134:333–344. - PubMed
-
- Fan, A.C.Y., M.K. Bhangoo, and J.C. Yound. 2006. Hsp90 functions in the targeting and outer membrane translocation steps of Tom70-mediated mitochondrial import. J. Biol. Chem. 281:33313–33324. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
