

Sequential roles for myosin-X in BMP6-dependent filopodial extension, migration, and activation of BMP receptors
- PMID: 18158328
- PMCID: PMC2373493
- DOI: 10.1083/jcb.200704010
Sequential roles for myosin-X in BMP6-dependent filopodial extension, migration, and activation of BMP receptors
Erratum in
- J Cell Biol. 2009 Feb 23;184(4):611
Abstract
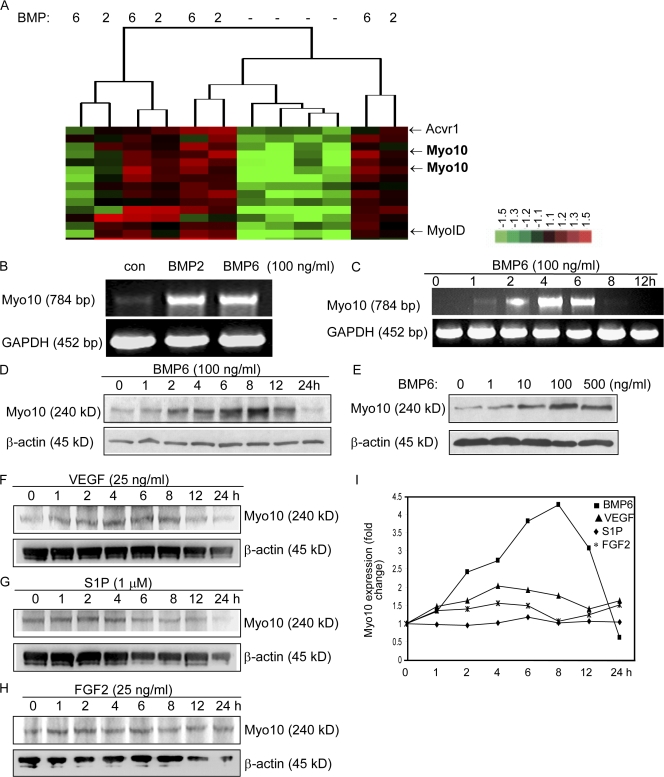
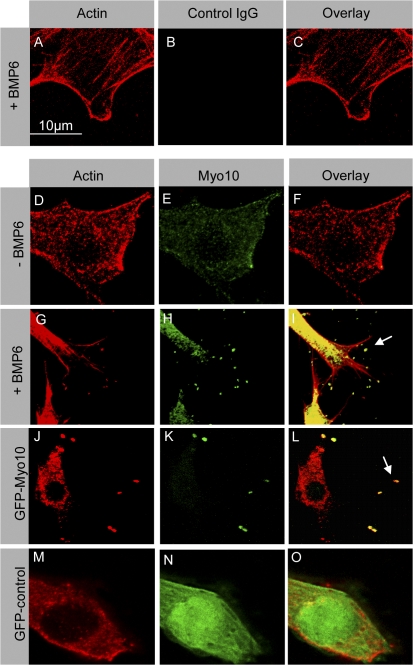
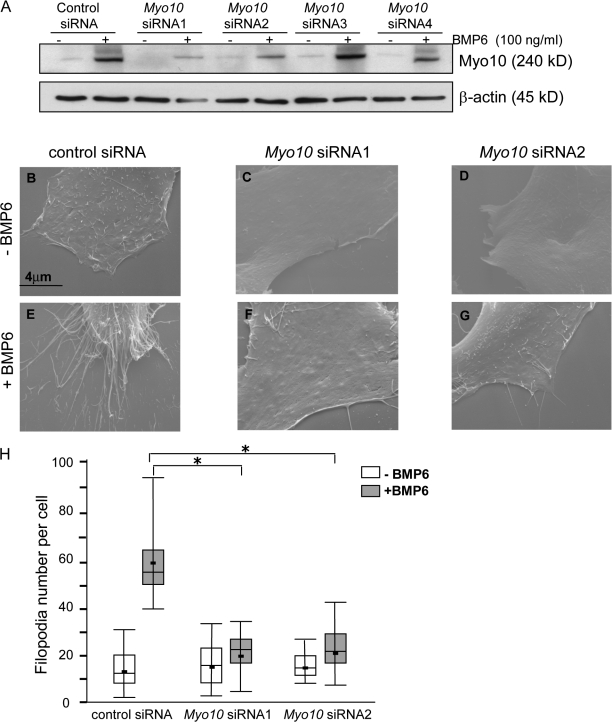
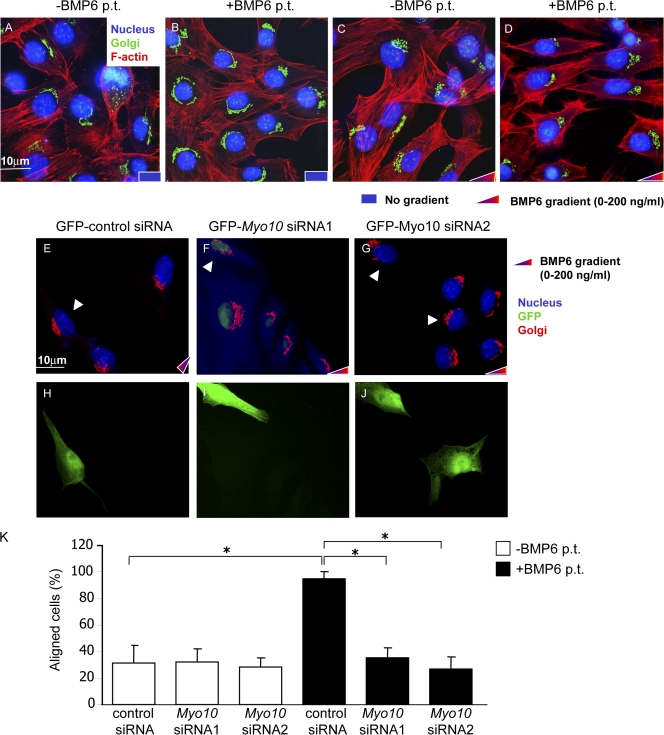
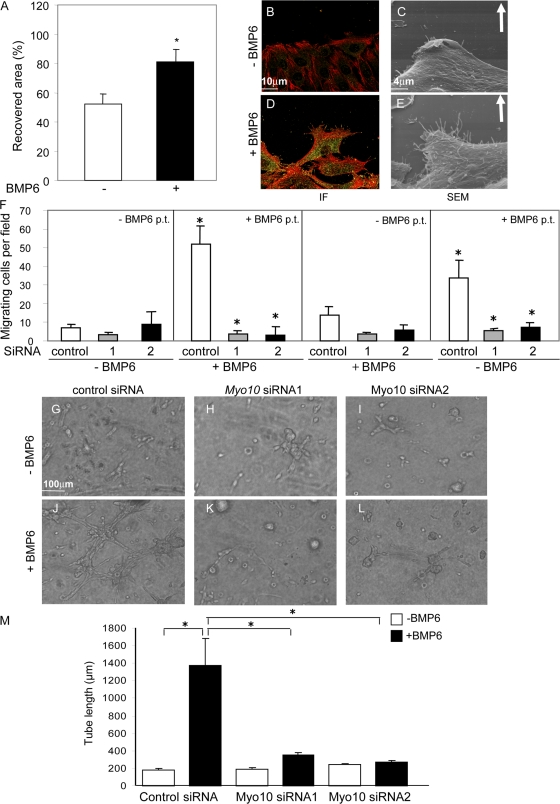
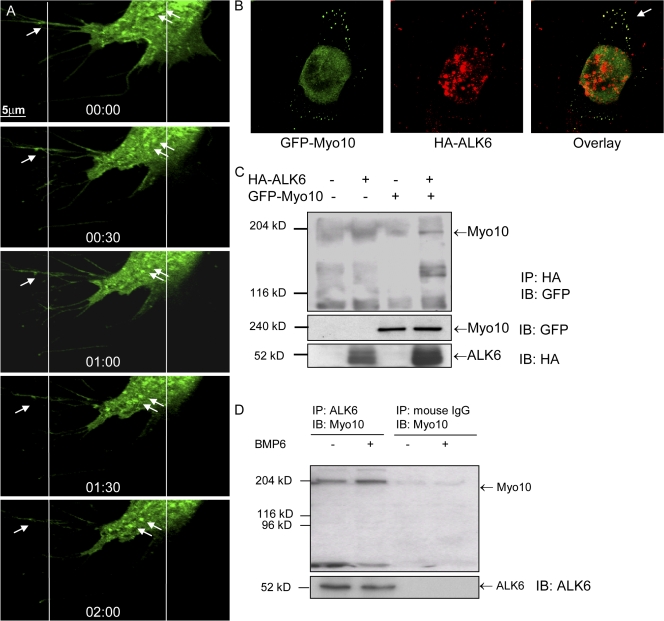
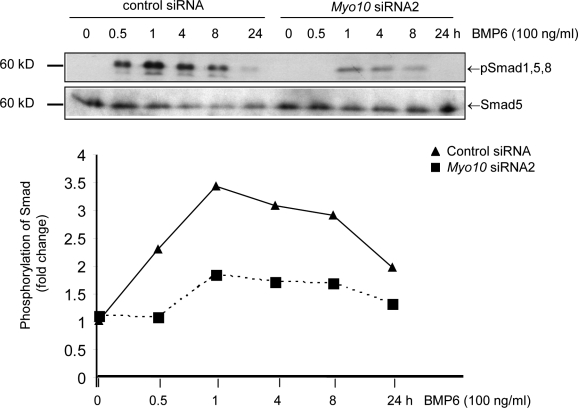
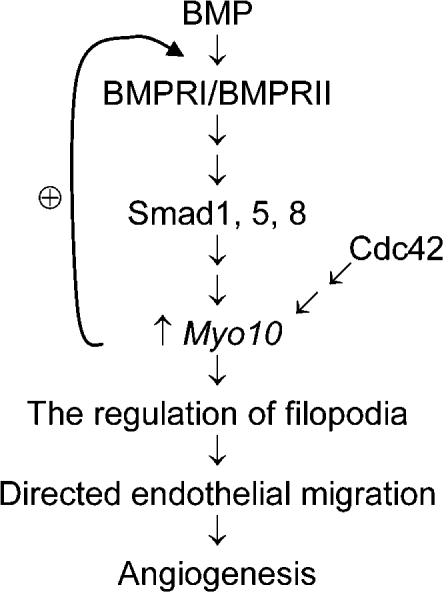
Endothelial cell migration is an important step during angiogenesis, and its dysregulation contributes to aberrant neovascularization. The bone morphogenetic proteins (BMPs) are potent stimulators of cell migration and angiogenesis. Using microarray analyses, we find that myosin-X (Myo10) is a BMP target gene. In endothelial cells, BMP6-induced Myo10 localizes in filopodia, and BMP-dependent filopodial assembly decreases when Myo10 expression is reduced. Likewise, cellular alignment and directional migration induced by BMP6 are Myo10 dependent. Surprisingly, we find that Myo10 and BMP6 receptor ALK6 colocalize in a BMP6-dependent fashion. ALK6 translocates into filopodia after BMP6 stimulation, and both ALK6 and Myo10 possess intrafilopodial motility. Additionally, Myo10 is required for BMP6-dependent Smad activation, indicating that in addition to its function in filopodial assembly, Myo10 also participates in a requisite amplification loop for BMP signaling. Our data indicate that Myo10 is required to guide endothelial migration toward BMP6 gradients via the regulation of filopodial function and amplification of BMP signals.
Figures
References
-
- Bar, T., and J.R. Wolff. 1972. The formation of capillary basement membranes during internal vascularization of the rat's cerebral cortex. Z. Zellforsch. Mikrosk. Anat. 133:231–248. - PubMed
-
- Berg, J.S., and R.E. Cheney. 2002. Myosin-X is an unconventional myosin that undergoes intrafilopodial motility. Nat. Cell Biol. 4:246–250. - PubMed
-
- Deckers, M.M., R.L. van Bezooijen, G. van der Horst, J. Hoogendam, C. van Der Bent, S.E. Papapoulos, and C.W. Lowik. 2002. Bone morphogenetic proteins stimulate angiogenesis through osteoblast-derived vascular endothelial growth factor A. Endocrinology. 143:1545–1553. - PubMed
-
- Folkman, J., and M. Klagsbrun. 1987. Vascular physiology. A family of angiogenic peptides. Nature. 329:671–672. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
