

Axons of retinal ganglion cells are insulted in the optic nerve early in DBA/2J glaucoma
- PMID: 18158332
- PMCID: PMC2373494
- DOI: 10.1083/jcb.200706181
Axons of retinal ganglion cells are insulted in the optic nerve early in DBA/2J glaucoma
Abstract
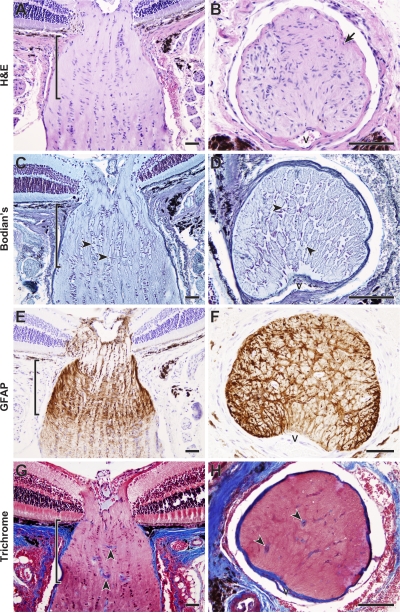
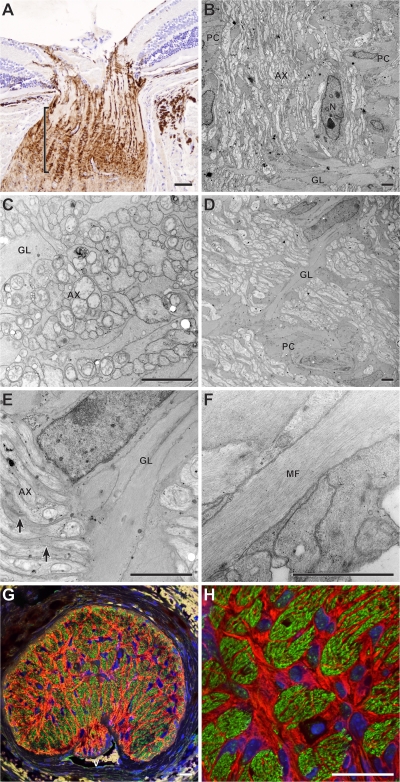
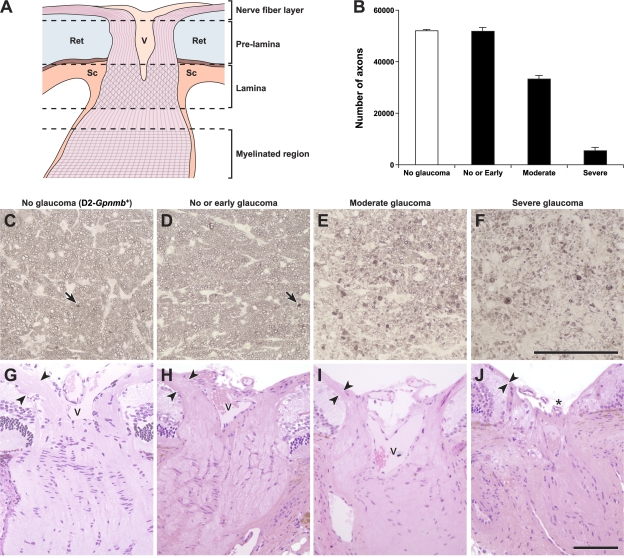
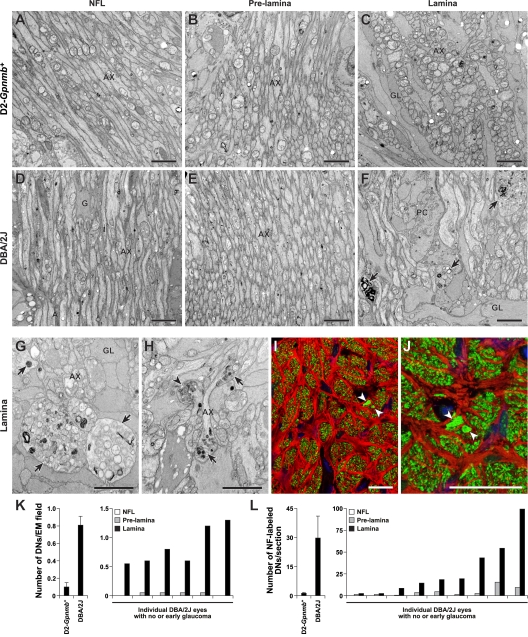
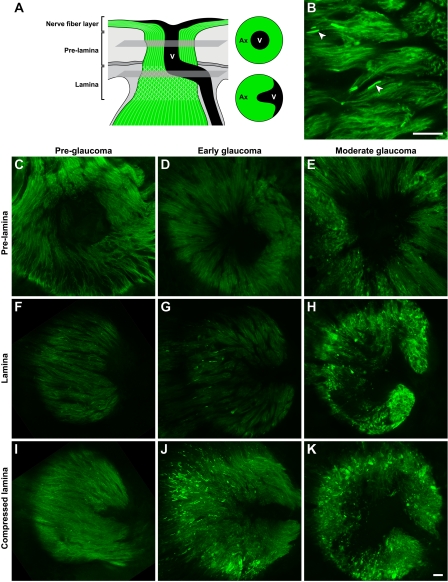
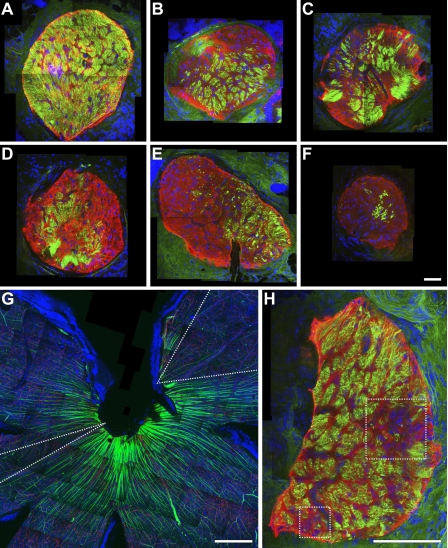
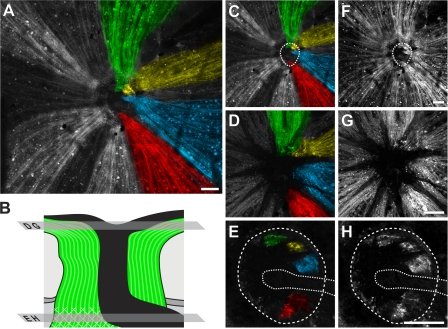
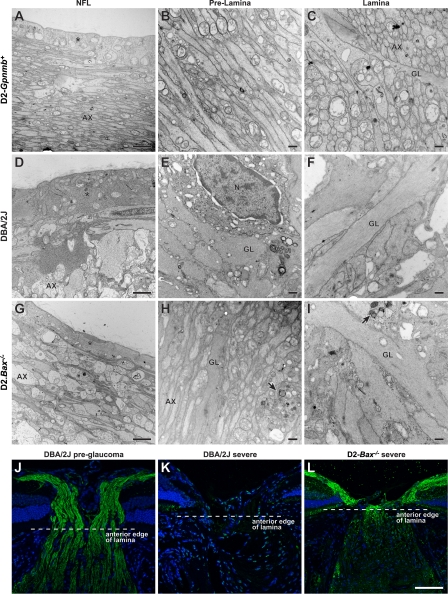
Here, we use a mouse model (DBA/2J) to readdress the location of insult(s) to retinal ganglion cells (RGCs) in glaucoma. We localize an early sign of axon damage to an astrocyte-rich region of the optic nerve just posterior to the retina, analogous to the lamina cribrosa. In this region, a network of astrocytes associates intimately with RGC axons. Using BAX-deficient DBA/2J mice, which retain all of their RGCs, we provide experimental evidence for an insult within or very close to the lamina in the optic nerve. We show that proximal axon segments attached to their cell bodies survive to the proximity of the lamina. In contrast, axon segments in the lamina and behind the eye degenerate. Finally, the Wld(s) allele, which is known to protect against insults to axons, strongly protects against DBA/2J glaucoma and preserves RGC activity as measured by pattern electroretinography. These experiments provide strong evidence for a local insult to axons in the optic nerve.
Figures

References
-
- Adalbert, R., T.H. Gillingwater, J.E. Haley, K. Bridge, B. Beirowski, L. Berek, D. Wagner, D. Grumme, D. Thomson, A. Celik, et al. 2005. A rat model of slow Wallerian degeneration (WldS) with improved preservation of neuromuscular synapses. Eur. J. Neurosci. 21:271–277. - PubMed
-
- Anderson, D.R. 1969. Ultrastructure of human and monkey lamina cribrosa and optic nerve head. Arch. Ophthalmol. 82:800–814. - PubMed
-
- Anderson, D.R. 1973. Ascending and descending optic atrophy produced experimentally in squirrel monkeys. Am. J. Ophthalmol. 76:693–711. - PubMed
-
- Anderson, D.R., and A. Hendrickson. 1974. Effect of intraocular pressure on rapid axoplasmic transport in monkey optic nerve. Invest. Ophthalmol. 13:771–783. - PubMed
-
- Anderson, D.R., and A.E. Hendrickson. 1977. Failure of increased intracranial pressure to affect rapid axonal transport at the optic nerve head. Invest. Ophthalmol. Vis. Sci. 16:423–426. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
