

Stress preconditioning of spreading depression in the locust CNS
- PMID: 18159249
- PMCID: PMC2137934
- DOI: 10.1371/journal.pone.0001366
Stress preconditioning of spreading depression in the locust CNS
Abstract
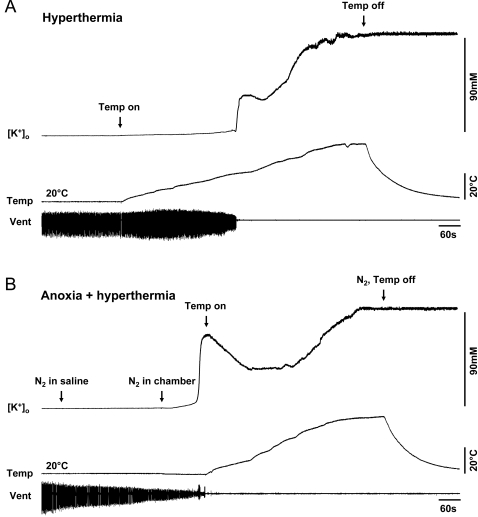
Cortical spreading depression (CSD) is closely associated with important pathologies including stroke, seizures and migraine. The mechanisms underlying SD in its various forms are still incompletely understood. Here we describe SD-like events in an invertebrate model, the ventilatory central pattern generator (CPG) of locusts. Using K(+) -sensitive microelectrodes, we measured extracellular K(+) concentration ([K(+)](o)) in the metathoracic neuropile of the CPG while monitoring CPG output electromyographically from muscle 161 in the second abdominal segment to investigate the role K(+) in failure of neural circuit operation induced by various stressors. Failure of ventilation in response to different stressors (hyperthermia, anoxia, ATP depletion, Na(+)/K(+) ATPase impairment, K(+) injection) was associated with a disturbance of CNS ion homeostasis that shares the characteristics of CSD and SD-like events in vertebrates. Hyperthermic failure was preconditioned by prior heat shock (3 h, 45 degrees C) and induced-thermotolerance was associated with an increase in the rate of clearance of extracellular K(+) that was not linked to changes in ATP levels or total Na(+)/K(+) ATPase activity. Our findings suggest that SD-like events in locusts are adaptive to terminate neural network operation and conserve energy during stress and that they can be preconditioned by experience. We propose that they share mechanisms with CSD in mammals suggesting a common evolutionary origin.
Conflict of interest statement
Figures






Similar articles
-
Pharmacological blockade of gap junctions induces repetitive surging of extracellular potassium within the locust CNS.J Insect Physiol. 2013 Oct;59(10):1031-40. doi: 10.1016/j.jinsphys.2013.07.007. Epub 2013 Aug 2. J Insect Physiol. 2013. PMID: 23916994
-
Suppression of spreading depression-like events in locusts by inhibition of the NO/cGMP/PKG pathway.J Neurosci. 2009 Jun 24;29(25):8225-35. doi: 10.1523/JNEUROSCI.1652-09.2009. J Neurosci. 2009. PMID: 19553462 Free PMC article.
-
Coma in response to environmental stress in the locust: a model for cortical spreading depression.J Insect Physiol. 2010 Aug;56(8):980-90. doi: 10.1016/j.jinsphys.2010.03.030. Epub 2010 Apr 7. J Insect Physiol. 2010. PMID: 20361971
-
Mechanisms of spreading depolarization in vertebrate and insect central nervous systems.J Neurophysiol. 2016 Sep 1;116(3):1117-27. doi: 10.1152/jn.00352.2016. Epub 2016 Jun 22. J Neurophysiol. 2016. PMID: 27334953 Free PMC article. Review.
-
A cold and quiet brain: mechanisms of insect CNS arrest at low temperatures.Curr Opin Insect Sci. 2023 Aug;58:101055. doi: 10.1016/j.cois.2023.101055. Epub 2023 May 16. Curr Opin Insect Sci. 2023. PMID: 37201631 Review.
Cited by
-
Neural shutdown under stress: an evolutionary perspective on spreading depolarization.J Neurophysiol. 2020 Mar 1;123(3):885-895. doi: 10.1152/jn.00724.2019. Epub 2020 Feb 5. J Neurophysiol. 2020. PMID: 32023142 Free PMC article. Review.
-
Alleviating brain stress: what alternative animal models have revealed about therapeutic targets for hypoxia and anoxia.Future Neurol. 2013;8(3):287-301. doi: 10.2217/fnl.13.12. Future Neurol. 2013. PMID: 25264428 Free PMC article.
-
Hyperkalaemia, not apoptosis, accurately predicts insect chilling injury.Proc Biol Sci. 2020 Dec 23;287(1941):20201663. doi: 10.1098/rspb.2020.1663. Epub 2020 Dec 16. Proc Biol Sci. 2020. PMID: 33323084 Free PMC article.
-
cGMP-Dependent Protein Kinase Inhibition Extends the Upper Temperature Limit of Stimulus-Evoked Calcium Responses in Motoneuronal Boutons of Drosophila melanogaster Larvae.PLoS One. 2016 Oct 6;11(10):e0164114. doi: 10.1371/journal.pone.0164114. eCollection 2016. PLoS One. 2016. PMID: 27711243 Free PMC article.
-
Impact of Heat Stress on Transposable Element Expression and Derived Small RNAs in Drosophila subobscura.Genome Biol Evol. 2023 Nov 1;15(11):evad189. doi: 10.1093/gbe/evad189. Genome Biol Evol. 2023. PMID: 37847062 Free PMC article.
References
-
- Wu J, Fisher RS. Hyperthermic spreading depressions in the immature rat hippocampal slice. J Neurophysiol. 2000;84:1355–1360. - PubMed
-
- Robertson RM. Modulation of neural circuit operation by prior environmental stress. Integr Comp Biol. 2004a;44:21–27. - PubMed
-
- Leão AAP. Spreading depression of activity in the cerebral cortex. J Neurophysiol. 1944;7:359–390. - PubMed
-
- Somjen GG. Mechanisms of spreading depression and hypoxic spreading depression-like depolarization. Physiol Rev. 2001;81:1065–1096. - PubMed
-
- Somjen GG. Ion regulation in the brain: implications for pathophysiology. Neuroscientist. 2002;8:254–267. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
