

Review
doi: 10.1523/JNEUROSCI.3578-07.2007.
Targeting and readout strategies for fast optical neural control in vitro and in vivo
Affiliations
- PMID: 18160630
- PMCID: PMC6673457
- DOI: 10.1523/JNEUROSCI.3578-07.2007
Item in Clipboard
Review
Targeting and readout strategies for fast optical neural control in vitro and in vivo
J Neurosci.
.
No abstract available
Figures
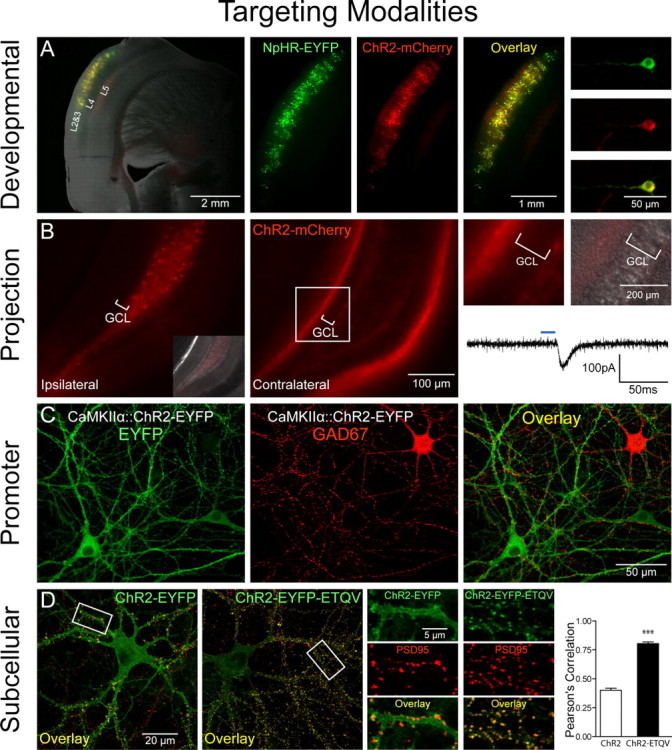
Targeting modalities. A, Developmental targeting: NpHR-EYFP (green) and ChR2-mCherry (red) expression in layers 2 and 3 of the mouse cortex via timed in utero electroporation (E16 surgery, P27 coronal slicing). A mixture of CMV::NpHR-EYFP and CMV::ChR2-mCherry DNA (1 μg/μl each) was pressure injected unilaterally into the ventricle and electroporated [5 50 ms pulses, 1 Hz, 40 V (see also Petreanu et al., 2007)]. Shown are pictures of acute slices taken on a Leica (Bannockburn, IL) MZ16FA stereomicroscope. B, Anterograde projection-based targeting: unilateral delivery of CaMKIIα::ChR2-mCherry lentivirus (1.5 μl, 109 pfu/ml) to the adult mouse dentate hilus/CA3 (left, and inset showing differential interference contrast image superposition) results in axonal ChR2-mCherry expression in the contralateral hippocampus (middle), with no expression in the granule cell layer (GCL) on either side of the brain (right). Note contralateral ChR2-mCherry-positive axonal projections to the molecular layer and subgranular zone. Optical stimulation with 15 ms pulses of blue light (HQ470 nm/40×; Chroma, Brattleboro, VT) of ChR2-positive axonal projections in acute slices from the contralateral hippocampus evoked inward synaptic currents in ChR2-negative dentate granule cells. A Lambda DG-4 optical switch (Sutter Instruments, Novato, CA) and 300 W xenon lamp were used for light delivery. Two injection sites were used at the following coordinates: anteroposterior (AP), 2 mm; mediolateral (ML), 1.5 mm; dorsoventral (DV), −2 mm; and AP, 3 mm; ML, 3 mm; DV, −3 mm. C, Promoter-based targeting: ChR2-EYFP (green) was targeted to excitatory hippocampal neurons in culture via CaMKIIα::ChR2-EYFP lentivirus [infected 2 d in vitro (div)]; immunostaining for GAD67 (red; 1:250; Millipore, Billerica, MA) in 14 div cultures shows exclusion of ChR2 expression from inhibitory GAD67+ neurons. D, Subcellular targeting: ChR2-EYFP (green) is expressed ubiquitously throughout the cell membrane in 16 div cultures. Adding the PDZ-domain binding motif ETQV to the C terminus by in-frame cloning led to concentration of ChR2-EYFP at postsynaptic sites, indicated by colocalization with PSD-95 staining (red; 1:200; Affinity Bioreagents, Golden, CO). Pearson's correlation (Manders et al., 1993) was r = 0.78 for ChR2-EYFP-ETQV and r = 0.40 for untargeted ChR2-EYFP; unpaired t test, ***p < 0.0001; n = 22 and 24 dendrites, respectively. Single confocal sections through 50 μm segments of proximal dendrites were thresholded and correlation coefficients determined using Volocity software.
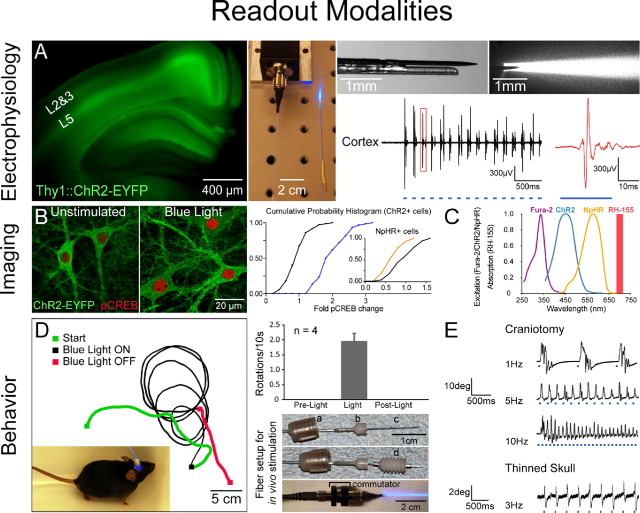
Readout modalities in vitro and in vivo. A, Electrophysiology in vivo: simultaneous optical stimulation and electrical recording from cortex in an intact Thy1::ChR2-EYFP mouse using an extracellular tungsten electrode (1 MΩ, ∼125 μm outer diameter) glued to an optical fiber (∼200 μm outer diameter; ThorLabs, Newton, NJ); the ∼0.5 mm offset (top right) ensures illumination of recorded neurons. The top-right illumination cone picture was generated by placing the electrode and fiber ensemble into a 1-cm-thick block of 1% agar (electrode superficial as shown); the geometrical dispersion is greatly modulated by scattering in brain tissue as described previously (Aravanis et al., 2007). FC/PC-coupled fiber optic is shown (middle) coupled to a 473 nm laser diode from CrystaLaser (Reno, NV) through an FC/PC adapter. Thy1::ChR2-EYFP (line 18) mice (Arenkiel et al., 2007; H. Wang et al., 2007) have strong ChR2 expression in layer 5 of neocortex (left). Recordings in anesthetized mice showed reliable cortex activation with pulsed blue light (10 Hz/25 ms), without stimulation artifacts at either the onset or offset of the stimulus. pClamp 10 and Digidata 1322A board (Molecular Devices, Union City, CA) were used to collect data and generate pulsed light. The recorded signal was bandpass filtered at 300 Hz low/5 kHz high (1800 Microelectrode AC Amplifier, A-M Systems, Sequim, WA). B, Post hoc biochemical imaging modality. Cultured hippocampal neurons [13–14 d in vitro (div), transduced 2 div with ChR2-EYFP and NpHR-EYFP lentiviruses under the CaMKIIα promoter) were preincubated ∼2 h with d -AP5 (25 μm ) and NBQX (10 μm ) and then stimulated with light in the continued presence of d -AP5/NBQX. Blue light delivery (HQ470 nm/40×; Chroma) for 10 min at 10 Hz/25 ms to ChR2-expressing neurons significantly elevated Ser133 pCREB compared with sham-treated neurons (unpaired t test, p < 0.0001; n = 60 and 72 cells, respectively). Inset, Activation of NpHR with 10 min of continuous amber light (593 nm/20×; Semrock, Rochester, NY) significantly lowered pCREB levels compared with sham-treated neurons (unpaired t test, p < 0.0001; n = 80 and 75 cells, respectively). Cumulative probability histograms show nuclear pCREB intensity normalized to the sham condition mean. For quantification of pCREB immunofluorescence (1:500; Millipore), five confocal images [40×/1.3 numerical aperture (NA) oil] were acquired per stimulated region. Single optical sections through the nucleus were analyzed using Volocity software, and nuclear regions were defined by DAPI labeling. Imaging and quantification were conducted blind to treatment condition. Optical stimulation was conducted on an Olympus (Tokyo, Japan) IX71 inverted scope using a 20×/0.75 NA objective [7 mW/mm2 light intensity measured with Newport (Irvine, CA) 1815-C power meter]. C, Real-time activity imaging modality. ChR2 and NpHR action spectra are shown superimposed with fura-2–Ca2+ indicator excitation spectrum and RH-155 voltage indicator absorption band. Spectral separation allows for simultaneous all-optical control and readout. D, Directional control of mammalian locomotion with fiber-optic light delivery in freely moving animals. Blue light stimulation of the right secondary motor cortex (M2) in Thy1::ChR2-EYFP adult mice induced repetitive rotations to the left that terminated immediately with conclusion of optical stimulation (1.96 ± 0.24 rotations/10 s). Light was delivered from the 473 nm laser via optical fiber through a fiber guide cannula (Plastics One, Roanoke, VA) targeted to the following coordinates: anteroposterior, 1 mm; mediolateral, 0.5 mm; dorsoventral, −0.5 mm. Pulses (30 Hz/15 ms) were generated using a function generator (33220A; Agilent, Palo Alto, CA). Cranioplastic cement was used to stabilize the fiber guide, and a dummy cannula (a) was placed while animals were not being tested. To ensure stability of the fiber during testing in moving animals, an internal cannula adapter (b) was glued to the stripped optical fiber (c), which was inserted into the fiber guide cannula fixed to the skull (d) (see also Adamantidis et al., 2007). A custom aluminum rotating optical commutator (bottom; Doric Lenses, Québec, Canada) was used to release torsion in the fiber caused by the animal's rotation. E, Whisking control with LEDs in sedated Thy1::ChR2-EYFP animals. Optical excitation of vibrissal motor cortex was performed either through a small craniotomy (top traces) or through thinned skull (bottom trace) with a blue LED [LEDtronics, Torrance, CA; skull thinned ∼80% with standard drill as in Yoder and Kleinfeld (2002)]. Whisker deflections were measured as before (Aravanis et al., 2007); a 1 mg rare-earth magnetic particle (Magcraft, Vienna, VA) was attached to the vibrissa, and a magnetoresistive sensor (Honeywell, Morristown, NJ) was used to measure deflections.
References
-
- Aravanis AM, Wang LP, Zhang F, Meltzer LA, Mogri MZ, Schneider MB, Deisseroth K. An optical neural interface: in vivo control of rodent motor cortex with integrated fiberoptic and optogenetic technology. J Neural Eng. 2007;4:S143–156. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources