

Overexpression of AtMYB44 enhances stomatal closure to confer abiotic stress tolerance in transgenic Arabidopsis
- PMID: 18162593
- PMCID: PMC2245844
- DOI: 10.1104/pp.107.110981
Overexpression of AtMYB44 enhances stomatal closure to confer abiotic stress tolerance in transgenic Arabidopsis
Abstract
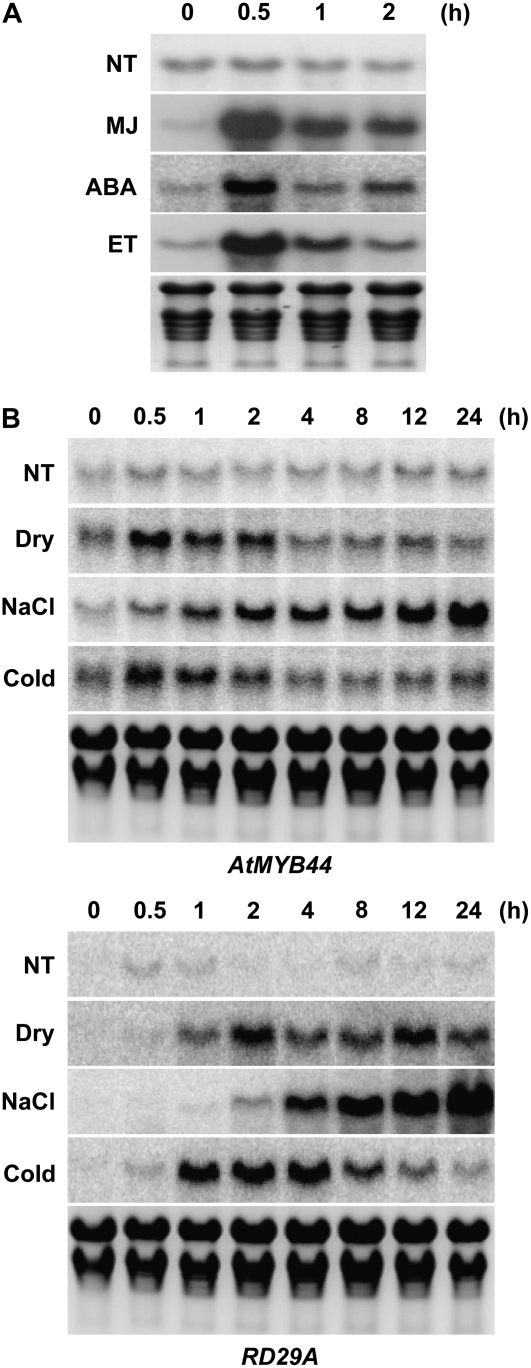
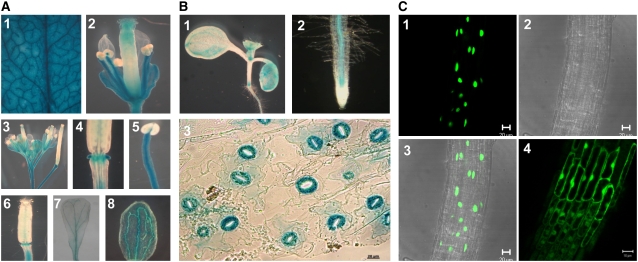
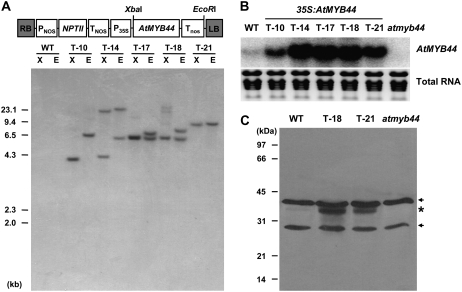
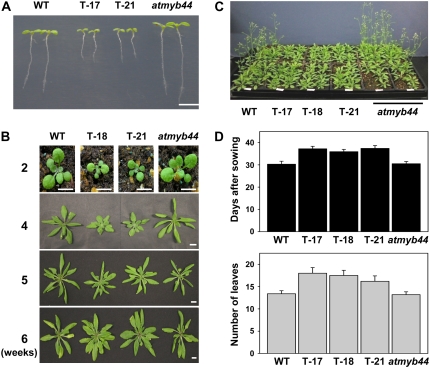
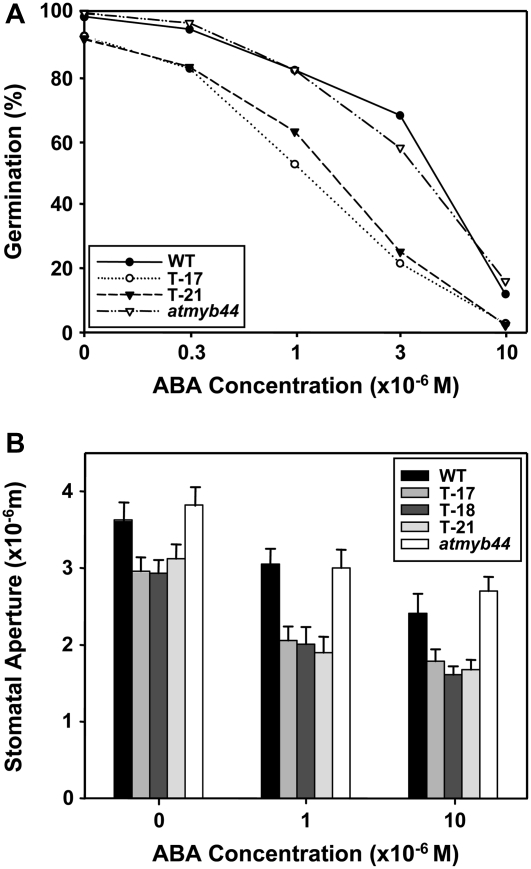
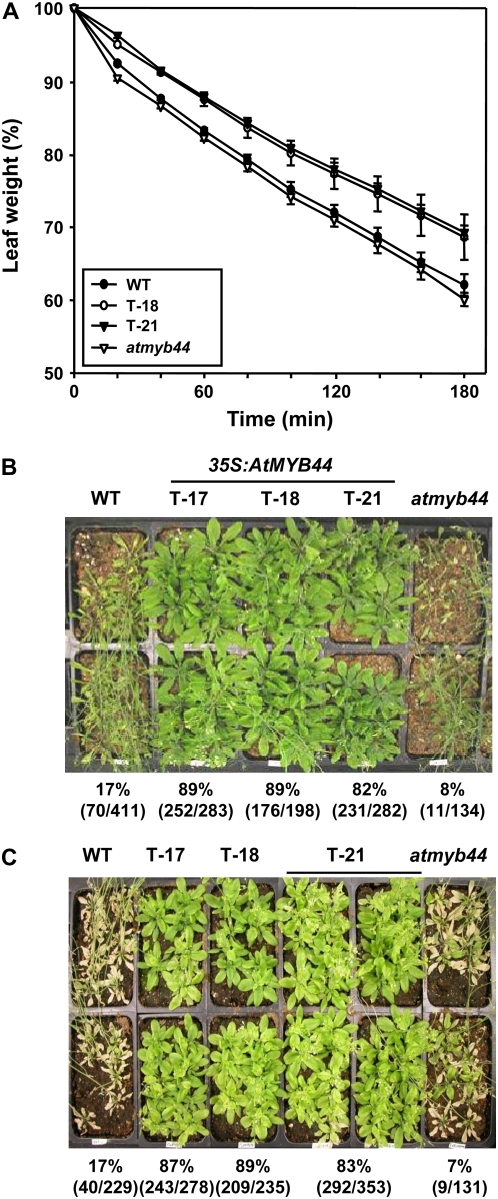
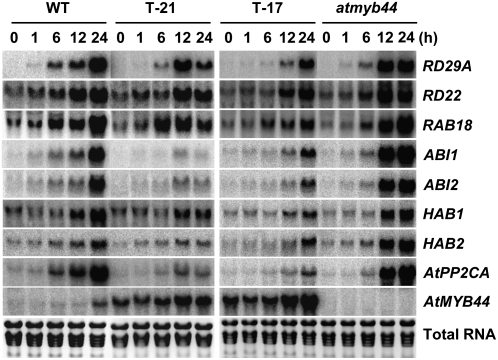
AtMYB44 belongs to the R2R3 MYB subgroup 22 transcription factor family in Arabidopsis (Arabidopsis thaliana). Treatment with abscisic acid (ABA) induced AtMYB44 transcript accumulation within 30 min. The gene was also activated under various abiotic stresses, such as dehydration, low temperature, and salinity. In transgenic Arabidopsis carrying an AtMYB44 promoter-driven beta-glucuronidase (GUS) construct, strong GUS activity was observed in the vasculature and leaf epidermal guard cells. Transgenic Arabidopsis overexpressing AtMYB44 is more sensitive to ABA and has a more rapid ABA-induced stomatal closure response than wild-type and atmyb44 knockout plants. Transgenic plants exhibited a reduced rate of water loss, as measured by the fresh-weight loss of detached shoots, and remarkably enhanced tolerance to drought and salt stress compared to wild-type plants. Microarray analysis and northern blots revealed that salt-induced activation of the genes that encode a group of serine/threonine protein phosphatases 2C (PP2Cs), such as ABI1, ABI2, AtPP2CA, HAB1, and HAB2, was diminished in transgenic plants overexpressing AtMYB44. By contrast, the atmyb44 knockout mutant line exhibited enhanced salt-induced expression of PP2C-encoding genes and reduced drought/salt stress tolerance compared to wild-type plants. Therefore, enhanced abiotic stress tolerance of transgenic Arabidopsis overexpressing AtMYB44 was conferred by reduced expression of genes encoding PP2Cs, which have been described as negative regulators of ABA signaling.
Figures
Similar articles
-
The FBA motif-containing protein AFBA1 acts as a novel positive regulator of ABA response in Arabidopsis.Plant Cell Physiol. 2017 Mar 1;58(3):574-586. doi: 10.1093/pcp/pcx003. Plant Cell Physiol. 2017. PMID: 28184867
-
Chromatin remodeling for the transcription of type 2C protein phosphatase genes in response to salt stress.Plant Physiol Biochem. 2019 Aug;141:325-331. doi: 10.1016/j.plaphy.2019.06.012. Epub 2019 Jun 11. Plant Physiol Biochem. 2019. PMID: 31207493
-
Transgenic expression of MYB15 confers enhanced sensitivity to abscisic acid and improved drought tolerance in Arabidopsis thaliana.J Genet Genomics. 2009 Jan;36(1):17-29. doi: 10.1016/S1673-8527(09)60003-5. J Genet Genomics. 2009. PMID: 19161942
-
Transgenic expression of rice MYB102 (OsMYB102) delays leaf senescence and decreases abiotic stress tolerance in Arabidopsis thaliana.BMB Rep. 2019 Nov;52(11):653-658. doi: 10.5483/BMBRep.2019.52.11.071. BMB Rep. 2019. PMID: 31072449 Free PMC article.
-
A stress-free walk from Arabidopsis to crops.Curr Opin Biotechnol. 2011 Apr;22(2):281-6. doi: 10.1016/j.copbio.2010.11.011. Epub 2010 Dec 17. Curr Opin Biotechnol. 2011. PMID: 21168324 Review.
Cited by
-
Lipid transfer protein 3 as a target of MYB96 mediates freezing and drought stress in Arabidopsis.J Exp Bot. 2013 Apr;64(6):1755-67. doi: 10.1093/jxb/ert040. Epub 2013 Feb 11. J Exp Bot. 2013. PMID: 23404903 Free PMC article.
-
MYB transcription factors-master regulators of phenylpropanoid biosynthesis and diverse developmental and stress responses.Plant Cell Rep. 2022 Dec;41(12):2245-2260. doi: 10.1007/s00299-022-02927-1. Epub 2022 Sep 29. Plant Cell Rep. 2022. PMID: 36171500 Review.
-
The Arabidopsis MIEL1 E3 ligase negatively regulates ABA signalling by promoting protein turnover of MYB96.Nat Commun. 2016 Sep 12;7:12525. doi: 10.1038/ncomms12525. Nat Commun. 2016. PMID: 27615387 Free PMC article.
-
CRISPR/Cas9 Gene Editing of NtAITRs, a Family of Transcription Repressor Genes, Leads to Enhanced Drought Tolerance in Tobacco.Int J Mol Sci. 2022 Dec 3;23(23):15268. doi: 10.3390/ijms232315268. Int J Mol Sci. 2022. PMID: 36499605 Free PMC article.
-
Expression of StMYB1R-1, a novel potato single MYB-like domain transcription factor, increases drought tolerance.Plant Physiol. 2011 Jan;155(1):421-32. doi: 10.1104/pp.110.163634. Epub 2010 Oct 27. Plant Physiol. 2011. PMID: 21030505 Free PMC article.
References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301 653–657 - PubMed
-
- Arabidopsis Genome Initiative (2000) Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408 796–815 - PubMed
-
- Carpenter CD, Simon AE (1998) Preparation of RNA. Methods Mol Biol 82 85–89 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
