

A spatially explicit model of muscle contraction explains a relationship between activation phase, power and ATP utilization in insect flight
- PMID: 18165245
- PMCID: PMC6902867
- DOI: 10.1242/jeb.013466
A spatially explicit model of muscle contraction explains a relationship between activation phase, power and ATP utilization in insect flight
Abstract
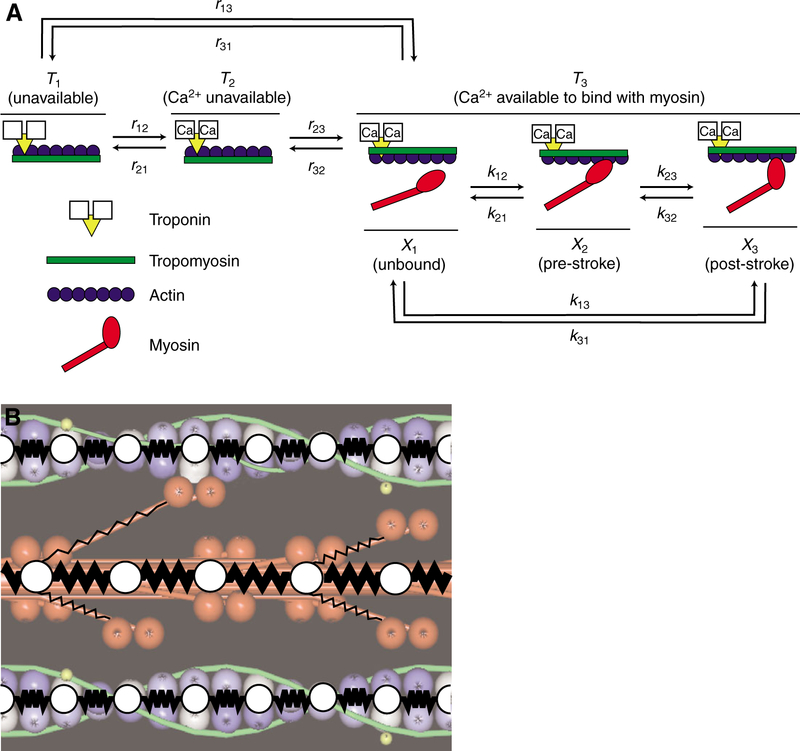
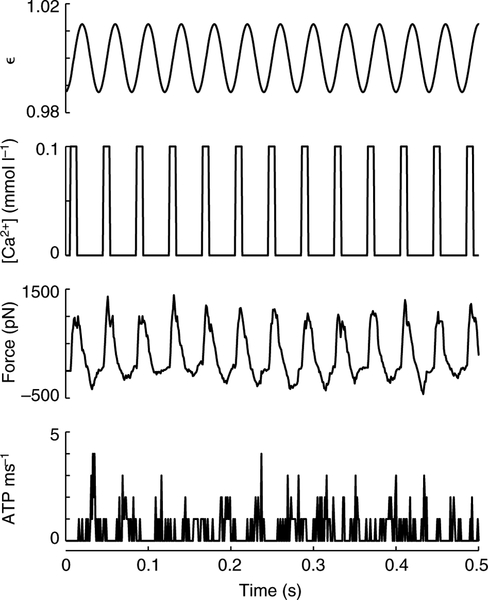
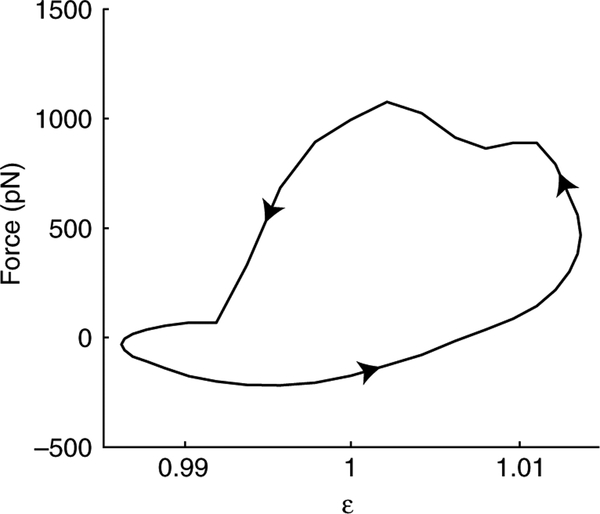
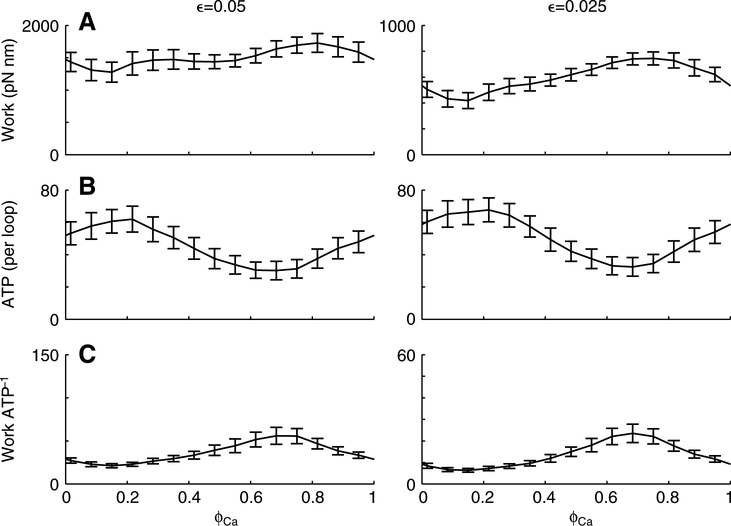
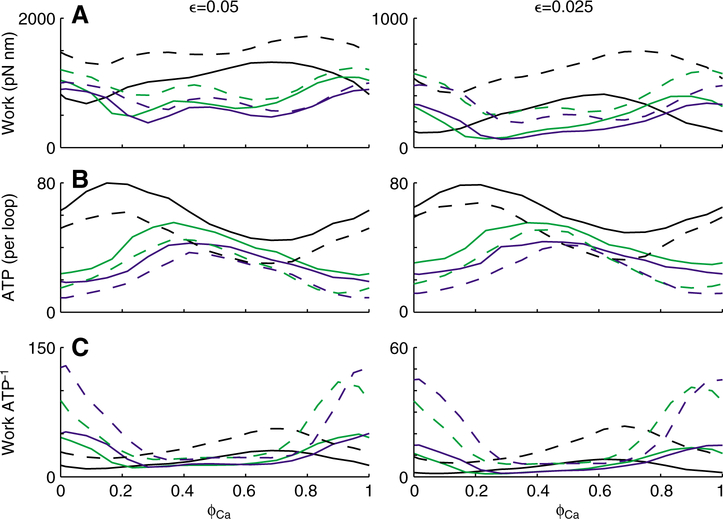
Using spatially explicit, stochastically kinetic, molecular models of muscle force generation, we examined the relationship between mechanical power output and energy utilization under differing patterns of length change and activation. A simulated work loop method was used to understand prior observations of sub-maximal power output in the dominant flight musculature of the hawkmoth Manduca sexta L. Here we show that mechanical work output and energy consumption (via ATP) vary with the phase of activation, although they do so with different phase sensitivities. The phase relationship for contraction efficiency (the ratio of power output to power input) differs from the phase relationships of energy consumption and power output. To our knowledge, this is the first report to suggest that ATP utilization by myosin cross-bridges varies strongly with the phase of activation in muscle undergoing cyclic length changes.
Figures
References
-
- Chase PB, MacPherson JM and Daniel TL (2004). A spatially explicit model of the half sarcomere: myofilament compliance affects Ca2+ regulation. Ann. Biomed. Eng 32, 1559–1568. - PubMed
-
- Curtin N and Woledge R (1996). Power at the expense of efficiency in contraction of white muscle fibres from dogfish Scyliorhinus canicula. J. Exp. Biol 199, 593–601. - PubMed
-
- Daniel TL and Tu MS (1999). Animal movement: mechanical tuning and coupled systems. J. Exp. Biol 202, 3415–3421. - PubMed
-
- Dickinson MH and Lighton JRB (1995). Muscle efficiency and elastic storage in the flight motor of Drosophila. Science 268, 87–90. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
