

Parasitism by Cuscuta pentagona attenuates host plant defenses against insect herbivores
- PMID: 18165323
- PMCID: PMC2259072
- DOI: 10.1104/pp.107.112219
Parasitism by Cuscuta pentagona attenuates host plant defenses against insect herbivores
Abstract
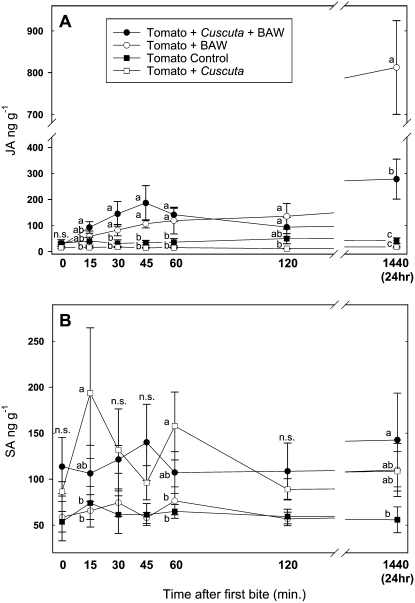
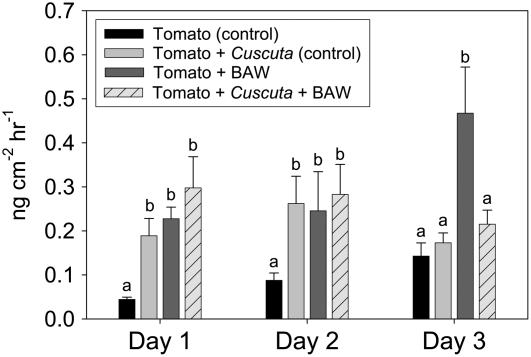
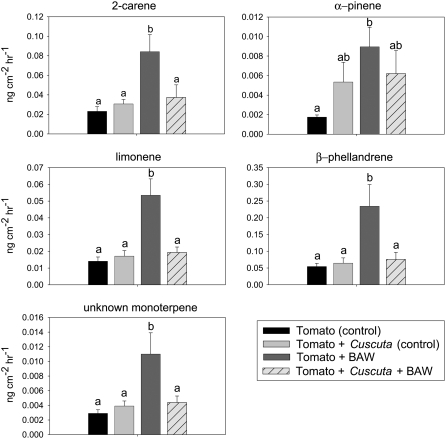
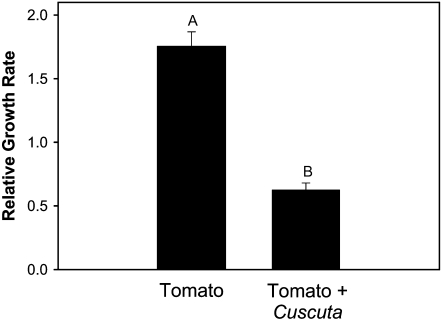
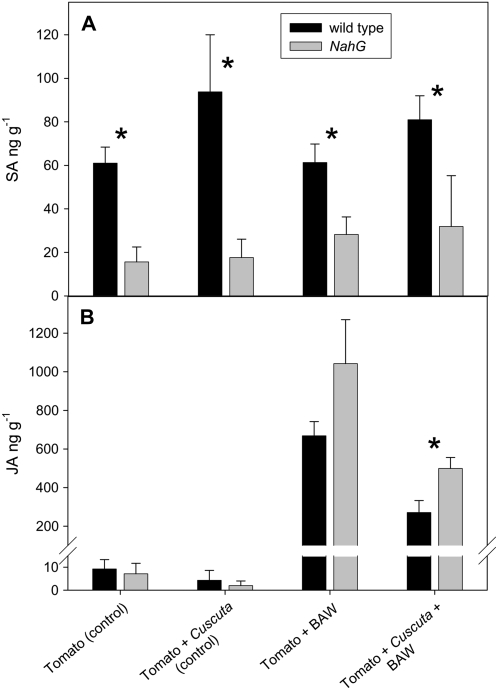
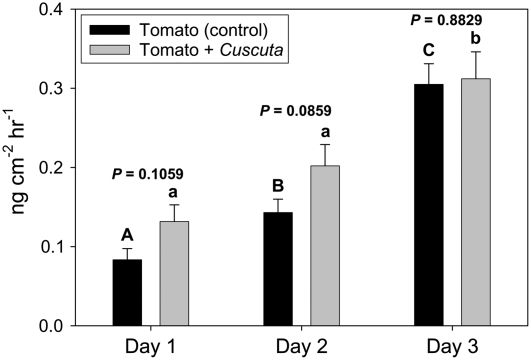
Considerable research has examined plant responses to concurrent attack by herbivores and pathogens, but the effects of attack by parasitic plants, another important class of plant-feeding organisms, on plant defenses against other enemies has not been explored. We investigated how attack by the parasitic plant Cuscuta pentagona impacted tomato (Solanum lycopersicum) defenses against the chewing insect beet armyworm (Spodoptera exigua; BAW). In response to insect feeding, C. pentagona-infested (parasitized) tomato plants produced only one-third of the antiherbivore phytohormone jasmonic acid (JA) produced by unparasitized plants. Similarly, parasitized tomato, in contrast to unparasitized plants, failed to emit herbivore-induced volatiles after 3 d of BAW feeding. Although parasitism impaired antiherbivore defenses, BAW growth was slower on parasitized tomato leaves. Vines of C. pentagona did not translocate JA from BAW-infested plants: amounts of JA in parasite vines grown on caterpillar-fed and control plants were similar. Parasitized plants generally contained more salicylic acid (SA), which can inhibit JA in some systems. Parasitized mutant (NahG) tomato plants deficient in SA produced more JA in response to insect feeding than parasitized wild-type plants, further suggesting cross talk between the SA and JA defense signaling pathways. However, JA induction by BAW was still reduced in parasitized compared to unparasitized NahG, implying that other factors must be involved. We found that parasitized plants were capable of producing induced volatiles when experimentally treated with JA, indicating that resource depletion by the parasite does not fully explain the observed attenuation of volatile response to herbivore feeding. Collectively, these findings show that parasitic plants can have important consequences for host plant defense against herbivores.
Figures
Similar articles
-
Plant defenses against parasitic plants show similarities to those induced by herbivores and pathogens.Plant Signal Behav. 2010 Aug;5(8):929-31. doi: 10.4161/psb.5.8.11772. Epub 2010 Aug 1. Plant Signal Behav. 2010. PMID: 20495380 Free PMC article.
-
Parasitism by Cuscuta pentagona sequentially induces JA and SA defence pathways in tomato.Plant Cell Environ. 2010 Feb;33(2):290-303. doi: 10.1111/j.1365-3040.2009.02082.x. Epub 2009 Nov 17. Plant Cell Environ. 2010. PMID: 19930126
-
Jasmonate-dependent induction of polyphenol oxidase activity in tomato foliage is important for defense against Spodoptera exigua but not against Manduca sexta.BMC Plant Biol. 2014 Sep 27;14:257. doi: 10.1186/s12870-014-0257-8. BMC Plant Biol. 2014. PMID: 25261073 Free PMC article.
-
Jasmonate- and salicylate-mediated plant defense responses to insect herbivores, pathogens and parasitic plants.Pest Manag Sci. 2009 May;65(5):497-503. doi: 10.1002/ps.1714. Pest Manag Sci. 2009. PMID: 19206090 Review.
-
Evolution of jasmonate and salicylate signal crosstalk.Trends Plant Sci. 2012 May;17(5):260-70. doi: 10.1016/j.tplants.2012.02.010. Epub 2012 Apr 11. Trends Plant Sci. 2012. PMID: 22498450 Review.
Cited by
-
Plant defenses against parasitic plants show similarities to those induced by herbivores and pathogens.Plant Signal Behav. 2010 Aug;5(8):929-31. doi: 10.4161/psb.5.8.11772. Epub 2010 Aug 1. Plant Signal Behav. 2010. PMID: 20495380 Free PMC article.
-
Messages from the Other Side: Parasites Receive Damage Cues from their Host Plants.J Chem Ecol. 2016 Aug;42(8):821-828. doi: 10.1007/s10886-016-0746-3. Epub 2016 Aug 18. J Chem Ecol. 2016. PMID: 27539588
-
Herbivory by the insect diaphorina citri induces greater change in citrus plant volatile profile than does infection by the bacterium, Candidatus Liberibacter asiaticus.Plant Signal Behav. 2013 Oct;8(10):doi: 10.4161/psb.25677. doi: 10.4161/psb.25677. Plant Signal Behav. 2013. PMID: 23857364 Free PMC article.
-
Parasite Removal, but Not Herbivory, Deters Future Parasite Attachment on Tomato.PLoS One. 2016 Aug 16;11(8):e0161076. doi: 10.1371/journal.pone.0161076. eCollection 2016. PLoS One. 2016. PMID: 27529694 Free PMC article.
-
Resource availability and parasitism intensity influence the response of soybean to the parasitic plant Cuscuta australis.Front Plant Sci. 2023 May 9;14:1177154. doi: 10.3389/fpls.2023.1177154. eCollection 2023. Front Plant Sci. 2023. PMID: 37229133 Free PMC article.
References
-
- Bardgett RD, Smith RS, Shiel RS, Peacock S, Simkin JM, Quirk H, Hobbs PJ (2006) Parasitic plants indirectly regulate below-ground properties in grassland ecosystems. Nature 439 969–972 - PubMed
-
- Birschwilks M, Sauer N, Scheel D, Neumann S (2007) Arabidopsis thaliana is a susceptible host plant for the holoparasite Cuscuta spec. Planta 226 1231–1241 - PubMed
-
- Borsics T, Lados M (2002) Dodder infection induces the expression of a pathogenesis-related gene of the family PR-10 in alfalfa. J Exp Bot 53 1831–1832 - PubMed
-
- Brading PA, Hammond-Kosack KE, Parr A, Jones JDG (2000) Salicylic acid is not required for Cf-2- and Cf-9-dependent resistance of tomato to Cladosporium fulvum. Plant J 23 305–318 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
