

The OSU1/QUA2/TSD2-encoded putative methyltransferase is a critical modulator of carbon and nitrogen nutrient balance response in Arabidopsis
- PMID: 18167546
- PMCID: PMC2148111
- DOI: 10.1371/journal.pone.0001387
The OSU1/QUA2/TSD2-encoded putative methyltransferase is a critical modulator of carbon and nitrogen nutrient balance response in Arabidopsis
Abstract
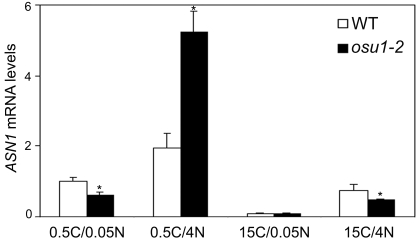
The balance between carbon (C) and nitrogen (N) nutrients must be tightly coordinated so that cells can optimize their opportunity for metabolism, growth and development. However, the C and N nutrient balance perception and signaling mechanism remains poorly understood. Here, we report the isolation and characterization of two allelic oversensitive to sugar 1 mutants (osu1-1, osu1-2) in Arabidopsis thaliana. Using the cotyledon anthocyanin accumulation and root growth inhibition assays, we show that the osu1 mutants are more sensitive than wild-type to both of the imbalanced C/N conditions, high C/low N and low C/high N. However, under the balanced C/N conditions (low C/low N or high C/high N), the osu1 mutants have similar anthocyanin levels and root lengths as wild-type. Consistently, the genes encoding two MYB transcription factors (MYB75 and MYB90) and an Asn synthetase isoform (ASN1) are strongly up-regulated by the OSU1 mutation in response to high C/low N and low C/high N, respectively. Furthermore, the enhanced sensitivity of osu1-1 to high C/low N with respect to anthocyanin accumulation but not root growth inhibition can be suppressed by co-suppression of MYB75, indicating that MYB75 acts downstream of OSU1 in the high C/low N imbalance response. Map-based cloning reveals that OSU1 encodes a member of a large family of putative methyltransferases and is allelic to the recently reported QUA2/TSD2 locus identified in genetic screens for cell-adhesion-defective mutants. Accumulation of OSU1/QUA2/TSD2 transcript was not regulated by C and N balance, but the OSU1 promoter was slightly more active in the vascular system. Taken together, our results show that the OSU1/QUA2/TSD2-encoded putative methyltransferase is required for normal C/N nutrient balance response in plants.
Conflict of interest statement
Figures






References
-
- Coruzzi GM, Zhou L. Carbon and nitrogen sensing and signaling in plants: emerging ‘matrix effects’. Curr Opin Plant Biol. 2001;4:247–253. - PubMed
-
- Commichau FM, Forchhammer K, Stulke J. Regulatory links between carbon and nitrogen metabolism. Curr Opin Microbiol. 2006;9:167–172. - PubMed
-
- Marshall S. Role of insulin, adipocyte hormones, and nutrient-sensing pathways in regulating fuel metabolism and energy homeostasis: a nutritional perspective of diabetes, obesity, and cancer. Sci STKE. 2006;2006:re7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
