

Phosphorylation of SNAP-25 at Ser187 mediates enhancement of exocytosis by a phorbol ester in INS-1 cells
- PMID: 18171919
- PMCID: PMC6671161
- DOI: 10.1523/JNEUROSCI.2352-07.2008
Phosphorylation of SNAP-25 at Ser187 mediates enhancement of exocytosis by a phorbol ester in INS-1 cells
Abstract
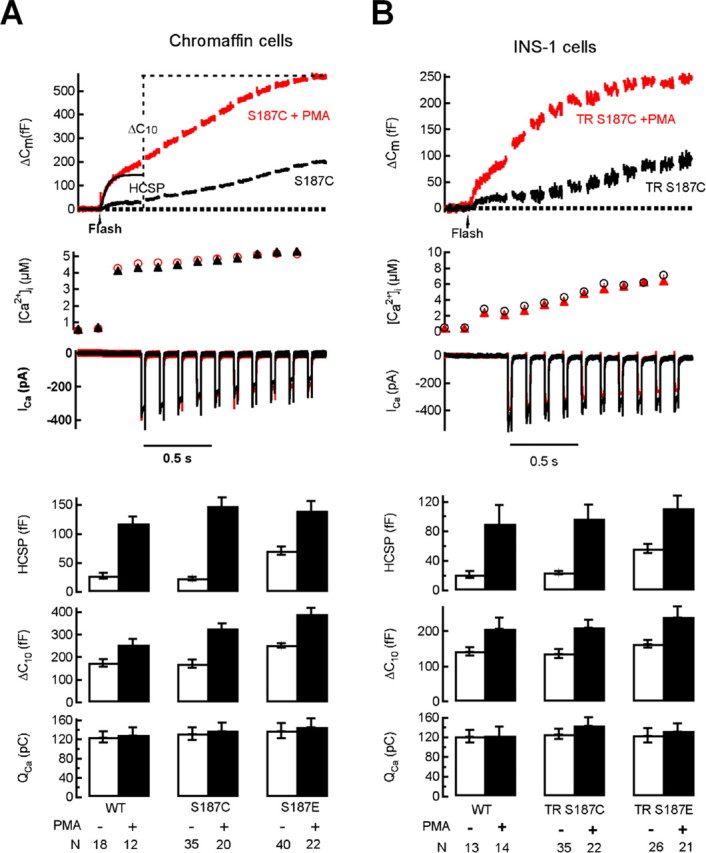
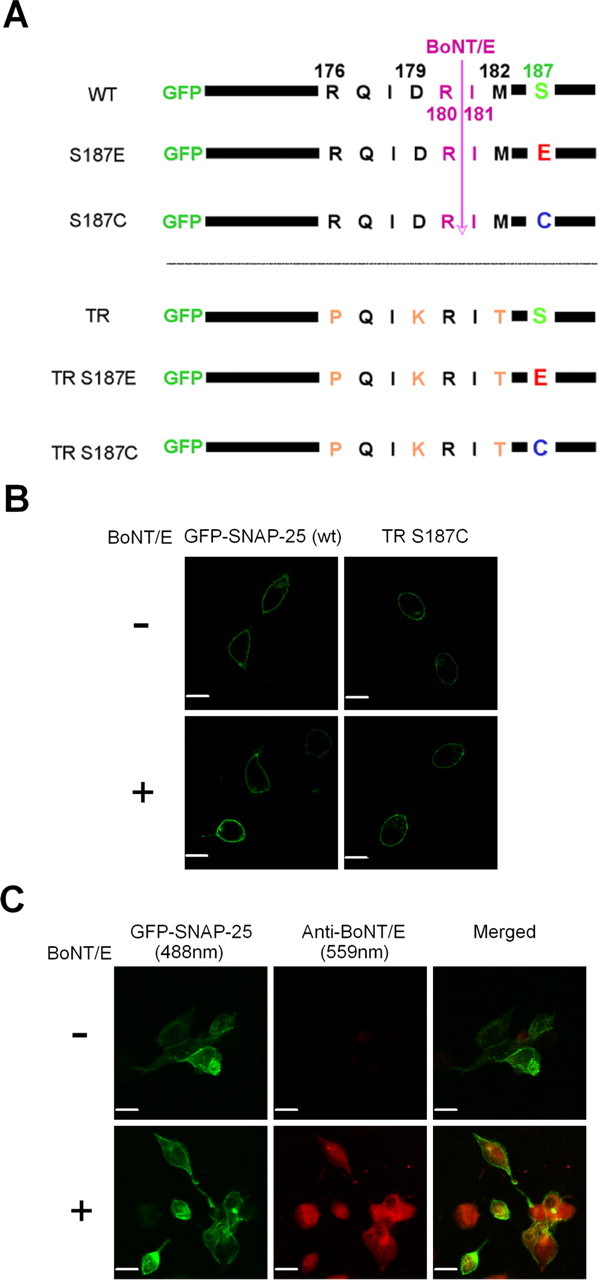
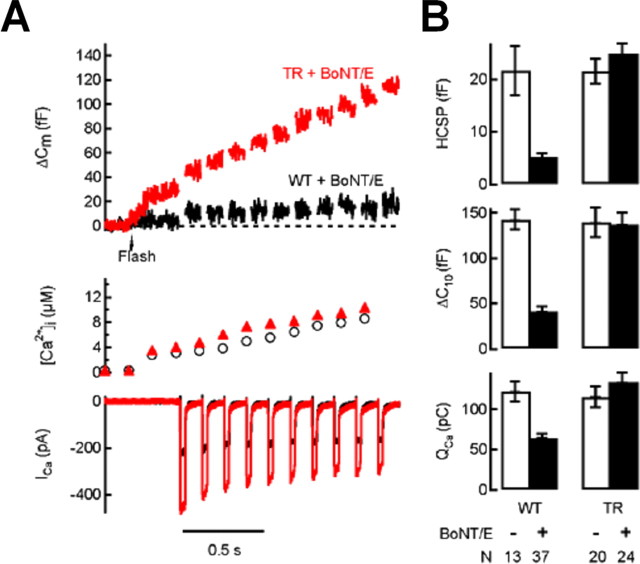
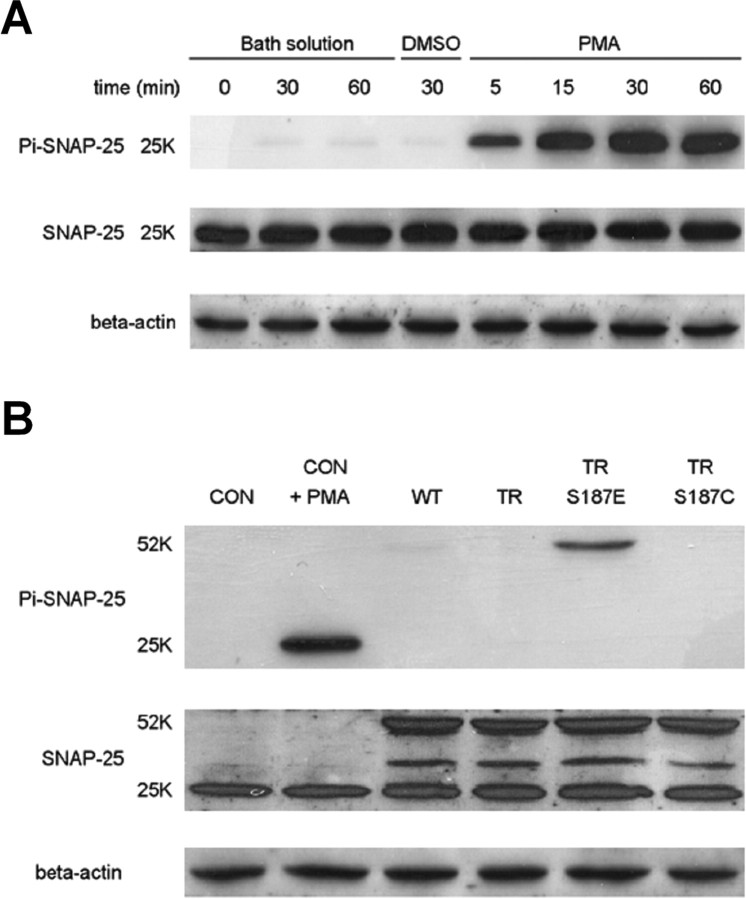
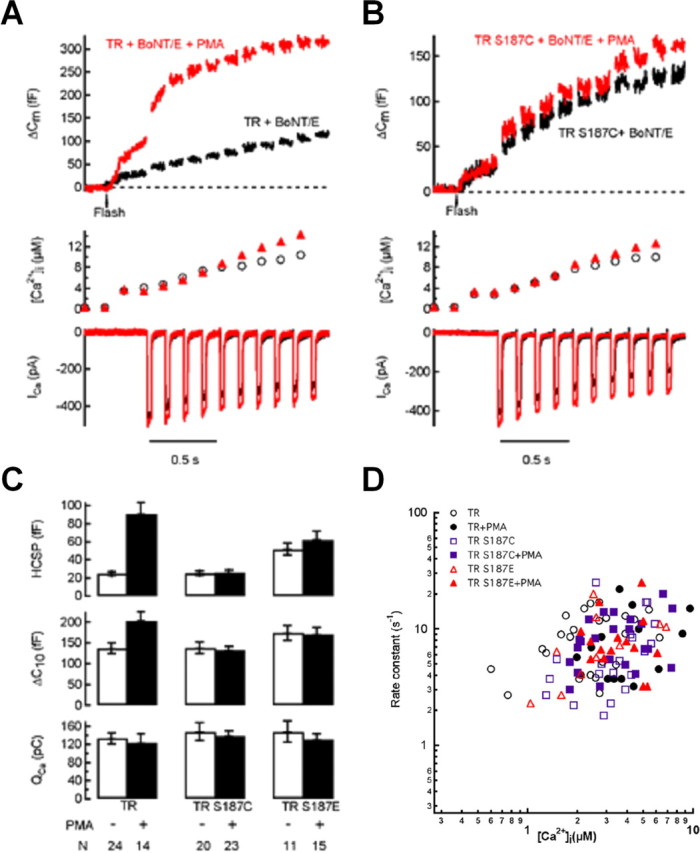
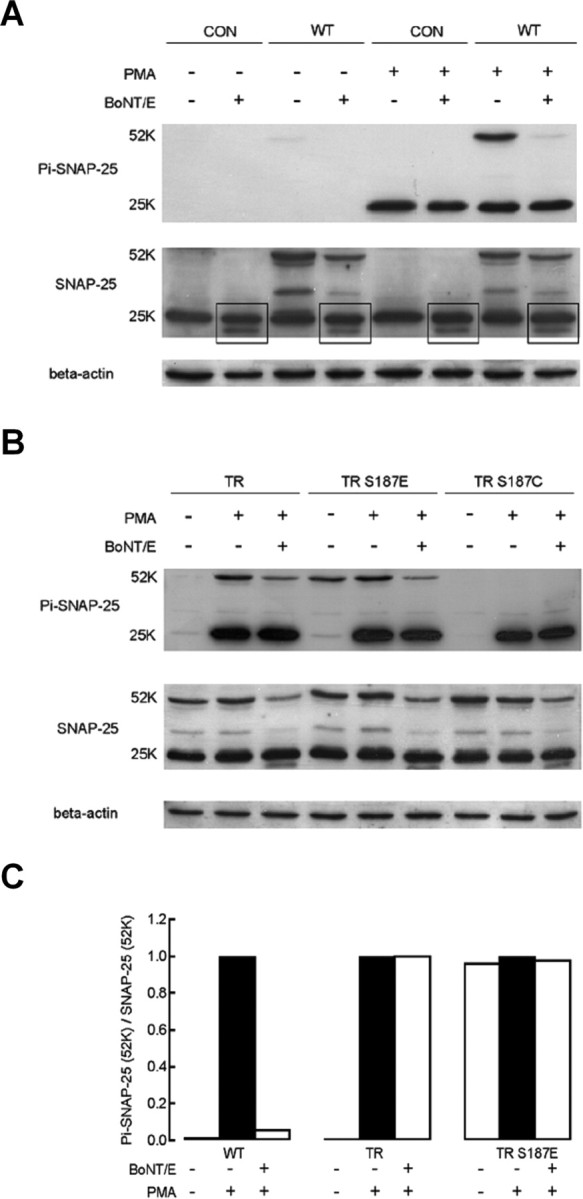
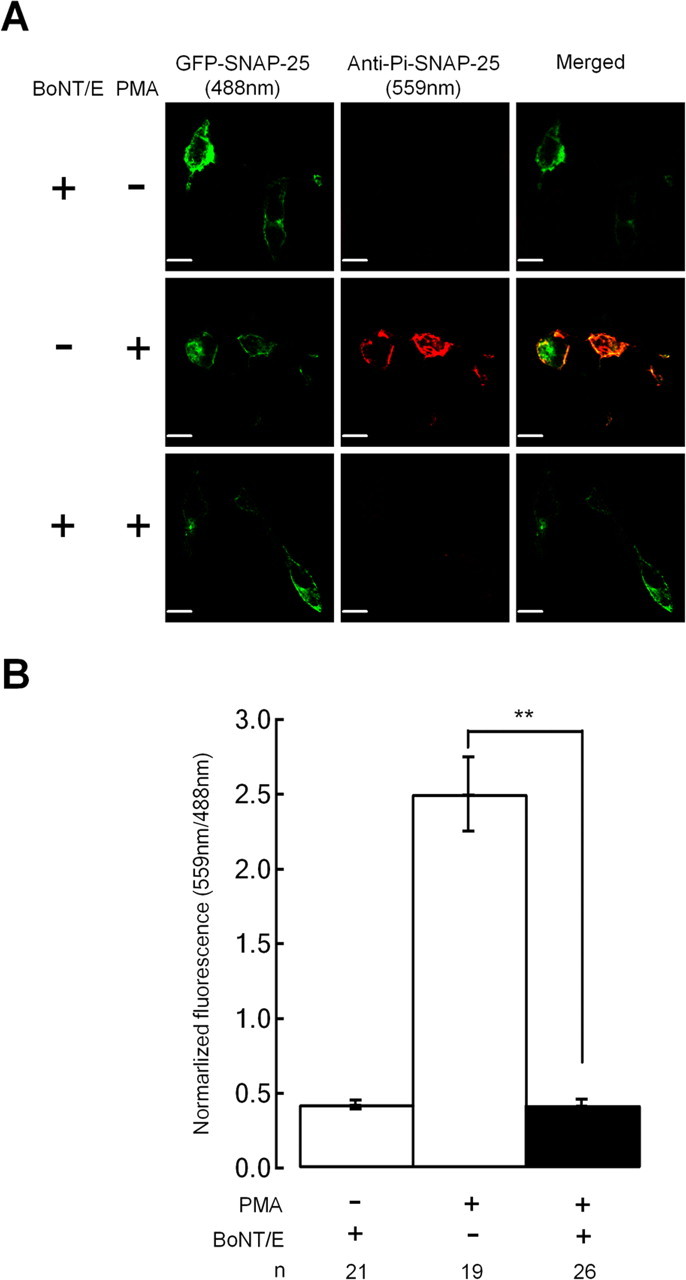
Activation of diacylglycerol (DAG) signaling pathways with phorbol esters dramatically enhances Ca2+-triggered exocytosis from both endocrine cells and neurons, however the relevant targets of DAG are controversial. A possible effector mechanism for this signaling pathway is phosphorylation of SNAP-25 (25 kDa synaptosome-associated protein) at Ser187 by PKC. Here, we investigated the role of Ser187 in the enhancement of exocytosis by the phorbol ester PMA (phorbol 12-myristate 13-acetate). We used patch-clamp measurements of membrane capacitance together with photorelease of caged-Ca2+ and membrane depolarization to study exocytosis. Expression of the nonphosphorylatable S187C SNAP-25 mutant did not attenuate the enhancement of exocytosis by PMA in either bovine chromaffin cells or the INS-1 insulin-secreting cell line. To test the effects of Ser187 mutations under conditions in which the endogenous SNAP-25 is disabled, we expressed botulinum toxin serotype E to cleave SNAP-25 in INS-1 cells. Coexpression of a toxin-resistant mutant (TR), but not wild-type SNAP-25, was able to rescue PMA-modulated exocytosis. Coexpression of the toxin with the TR-S187C SNAP-25 mutant was able to completely block the enhancement of exocytosis by PMA in response to photoelevation of [Ca2+]i to low microM levels or to a depolarizing train. The phospho-mimetic S187E mutation enhanced the small, fast burst of exocytosis evoked by photelevation of Ca2+, but, like PMA, had smaller effects on exocytosis evoked by a depolarizing train. This work supports the hypothesis that phosphorylation of Ser187 of SNAP-25 by PKC is a key step in the enhancement of exocytosis by DAG.
Figures
Similar articles
-
Phosphomimetic mutation of Ser-187 of SNAP-25 increases both syntaxin binding and highly Ca2+-sensitive exocytosis.J Gen Physiol. 2007 Mar;129(3):233-44. doi: 10.1085/jgp.200609685. J Gen Physiol. 2007. PMID: 17325194 Free PMC article.
-
SNAP-25 phosphorylation at Ser187 is not involved in Ca2+ or phorbolester-dependent potentiation of synaptic release.Mol Cell Neurosci. 2020 Jan;102:103452. doi: 10.1016/j.mcn.2019.103452. Epub 2019 Nov 30. Mol Cell Neurosci. 2020. PMID: 31794878
-
Phosphorylation of SNAP-25 on serine-187 is induced by secretagogues in insulin-secreting cells, but is not correlated with insulin secretion.Biochem J. 2002 Nov 15;368(Pt 1):223-32. doi: 10.1042/BJ20020896. Biochem J. 2002. PMID: 12164783 Free PMC article.
-
Protein kinase C-dependent phosphorylation of synaptosome-associated protein of 25 kDa at Ser187 potentiates vesicle recruitment.J Neurosci. 2002 Nov 1;22(21):9278-86. doi: 10.1523/JNEUROSCI.22-21-09278.2002. J Neurosci. 2002. PMID: 12417653 Free PMC article.
-
Regulation of exocytosis by protein kinase C.Biochem Soc Trans. 2005 Dec;33(Pt 6):1341-4. doi: 10.1042/BST0331341. Biochem Soc Trans. 2005. PMID: 16246114 Review.
Cited by
-
IκB kinase phosphorylation of SNAP-23 controls platelet secretion.Blood. 2013 May 30;121(22):4567-74. doi: 10.1182/blood-2012-11-470468. Epub 2013 Apr 23. Blood. 2013. PMID: 23613522 Free PMC article.
-
Modes of exocytosis and electrogenesis underlying canine biphasic insulin secretion.Front Biosci (Elite Ed). 2012 Jan 1;4(2):669-76. doi: 10.2741/e408. Front Biosci (Elite Ed). 2012. PMID: 22201903 Free PMC article. Review.
-
Single secretory granules of live cells recruit syntaxin-1 and synaptosomal associated protein 25 (SNAP-25) in large copy numbers.Proc Natl Acad Sci U S A. 2010 Nov 30;107(48):20810-5. doi: 10.1073/pnas.1014840107. Epub 2010 Nov 12. Proc Natl Acad Sci U S A. 2010. PMID: 21076040 Free PMC article.
-
A single amino acid mutation in SNAP-25 induces anxiety-related behavior in mouse.PLoS One. 2011;6(9):e25158. doi: 10.1371/journal.pone.0025158. Epub 2011 Sep 20. PLoS One. 2011. PMID: 21949876 Free PMC article.
-
Myosin phosphatase and RhoA-activated kinase modulate neurotransmitter release by regulating SNAP-25 of SNARE complex.PLoS One. 2017 May 9;12(5):e0177046. doi: 10.1371/journal.pone.0177046. eCollection 2017. PLoS One. 2017. PMID: 28486561 Free PMC article.
References
-
- Andres-Mateos E, Renart J, Cruces J, Solis-Garrido LM, Serantes R, de Lucas-Cerrillo AM, Aldea M, Garcia AG, Montiel C. Dynamic association of the Ca2+ channel alpha1A subunit and SNAP-25 in round or neurite-emitting chromaffin cells. Eur J Neurosci. 2005;22:2187–2198. - PubMed
-
- Asfari M, Janjic D, Meda P, Li G, Halban PA, Wollheim CB. Establishment of 2-mercaptoethanol-dependent differentiated insulin-secreting cell lines. Endocrinology. 1992;130:167–178. - PubMed
-
- Ashery U, Betz A, Xu T, Brose N, Rettig J. An efficient method for infection of adrenal chromaffin cells using the Semliki Forest virus gene expression system. Eur J Cell Biol. 1999;78:525–532. - PubMed
-
- Blasi J, Chapman ER, Link E, Binz T, Yamasaki S, De Camilli P, Sudhof TC, Niemann H, Jahn R. Botulinum neurotoxin A selectively cleaves the synaptic protein SNAP-25. Nature. 1993;365:160–163. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous