

Lrig1 is an endogenous inhibitor of Ret receptor tyrosine kinase activation, downstream signaling, and biological responses to GDNF
- PMID: 18171921
- PMCID: PMC6671136
- DOI: 10.1523/JNEUROSCI.2196-07.2008
Lrig1 is an endogenous inhibitor of Ret receptor tyrosine kinase activation, downstream signaling, and biological responses to GDNF
Abstract
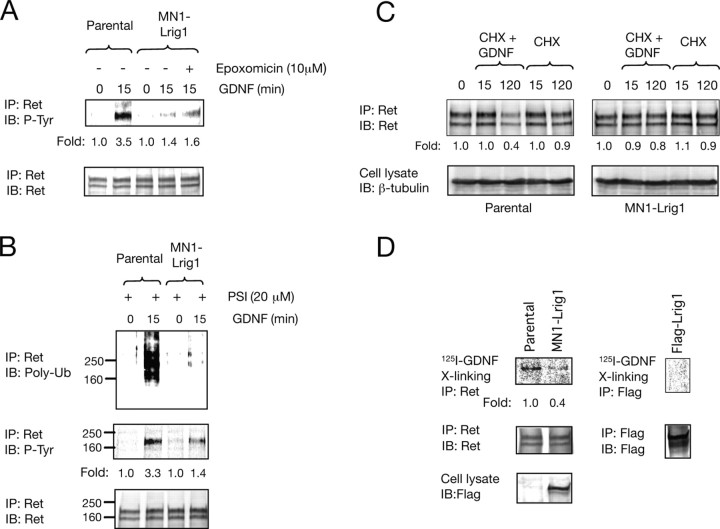
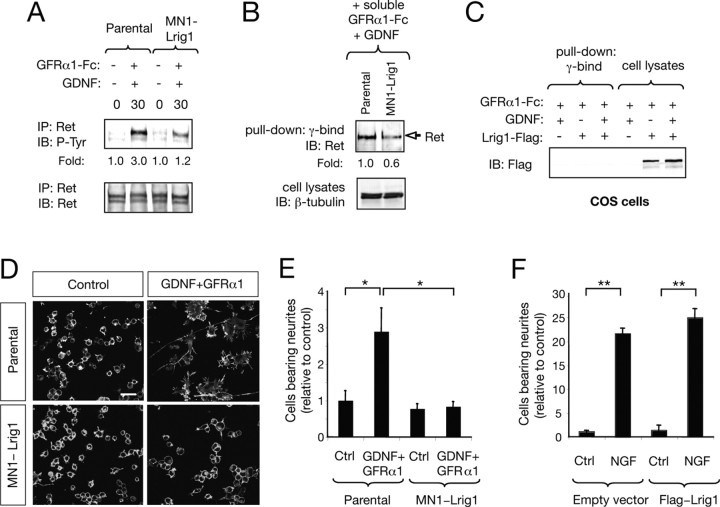
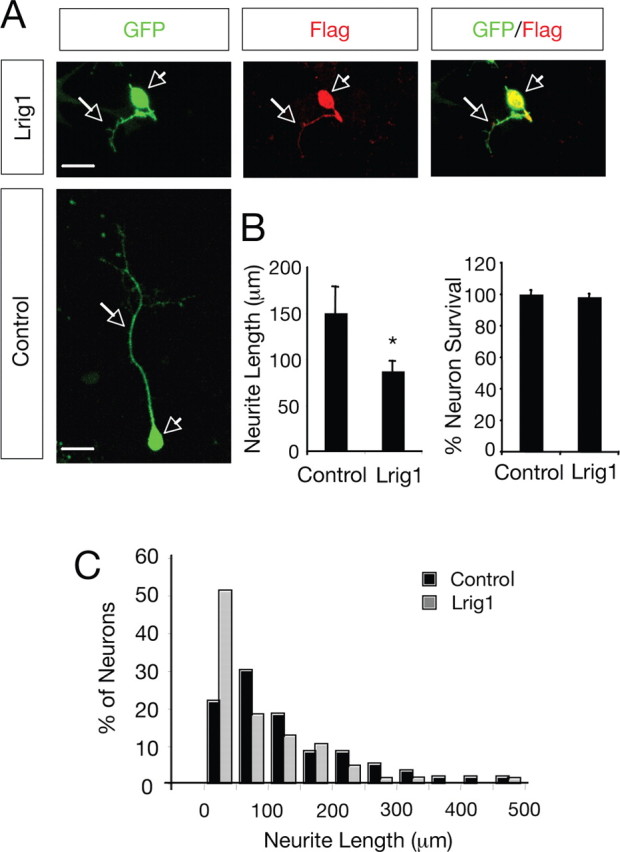
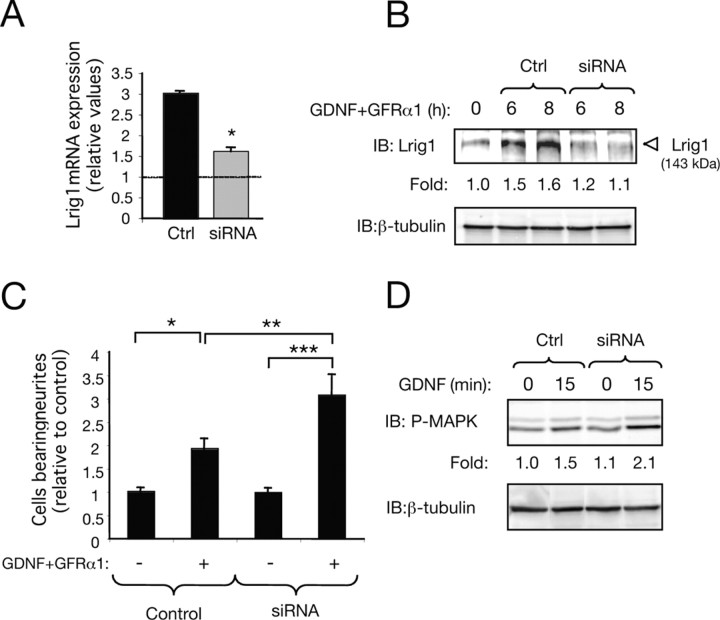
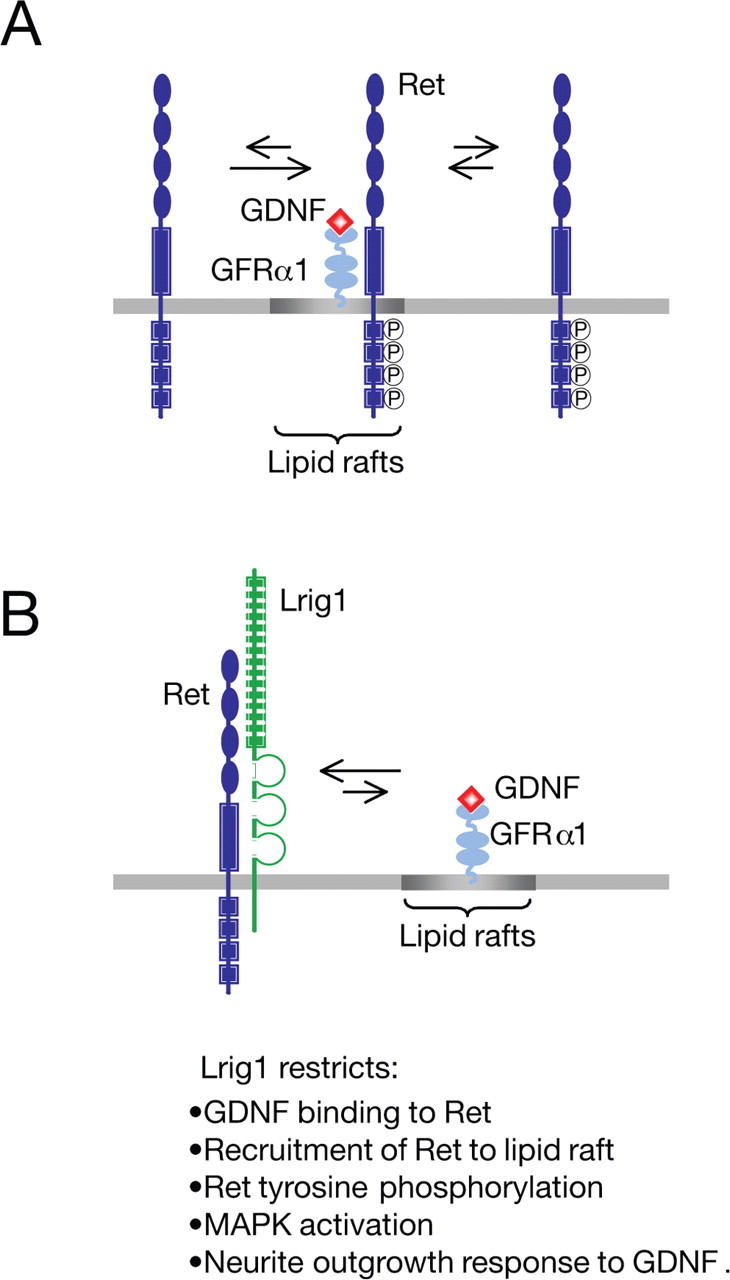
Glial cell line-derived neurotrophic factor (GDNF)/Ret signaling has potent trophic effects on ventral midbrain dopaminergic, motor, sensory, and sympathetic neurons. The molecular mechanisms that restrict Ret receptor tyrosine kinase activation are not well understood. Here, we show that Lrig1, a transmembrane protein containing leucine-rich repeats and Ig-like domains in its extracellular region, acts in a negative feedback loop to regulate the activity of Ret receptor tyrosine kinase. In particular, we demonstrate that Lrig1 is capable of physically interacting with Ret and that Lrig1/Ret association inhibits GDNF binding, recruitment of Ret to lipid rafts, receptor autophosphorylation, and mitogen-activated protein kinase (MAPK) activation in response to GDNF. In neuronal cells, Lrig1 overexpression also inhibits GDNF/Ret-induced neurite outgrowth in a cell-autonomous manner. Downregulation of Lrig1 using small interference RNA knock-down experiments potentiates both neuronal differentiation and MAPK activation in response to GDNF. Together, these results provide an insight into Lrig1 function and establish a new physiological mechanism to restrict signaling and biological responses induced by GDNF and Ret in neuronal cells.
Figures
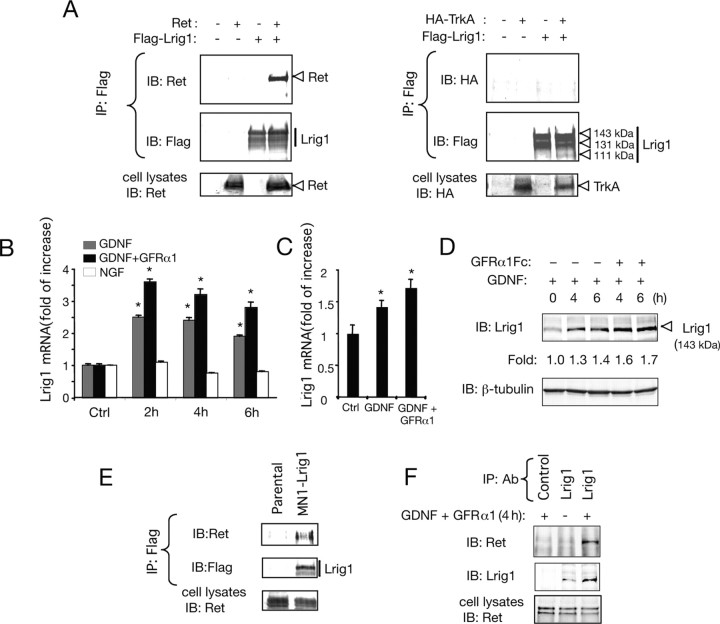
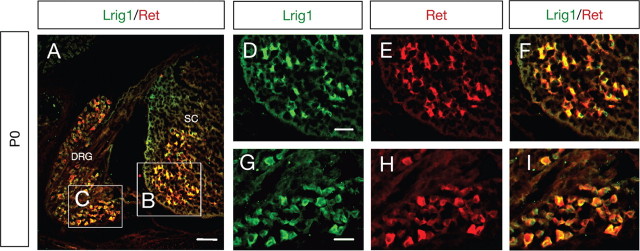
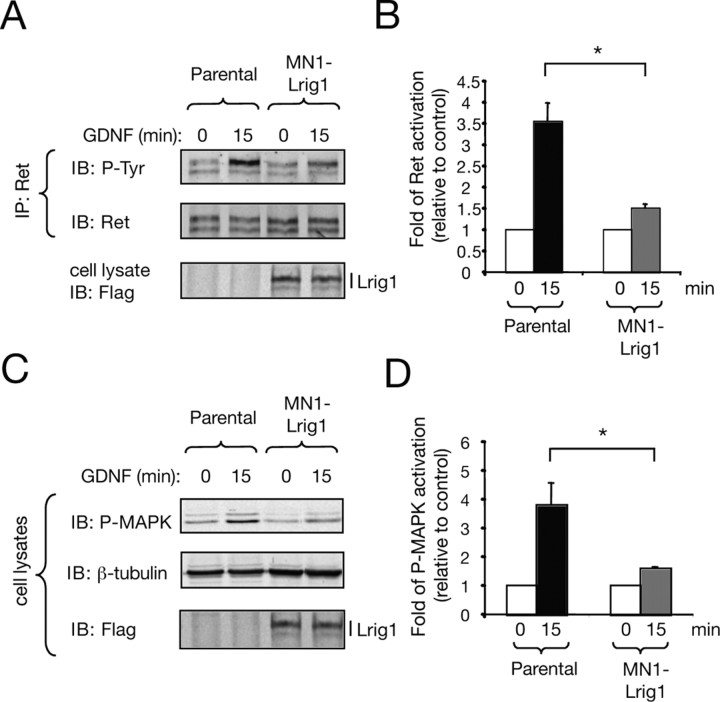
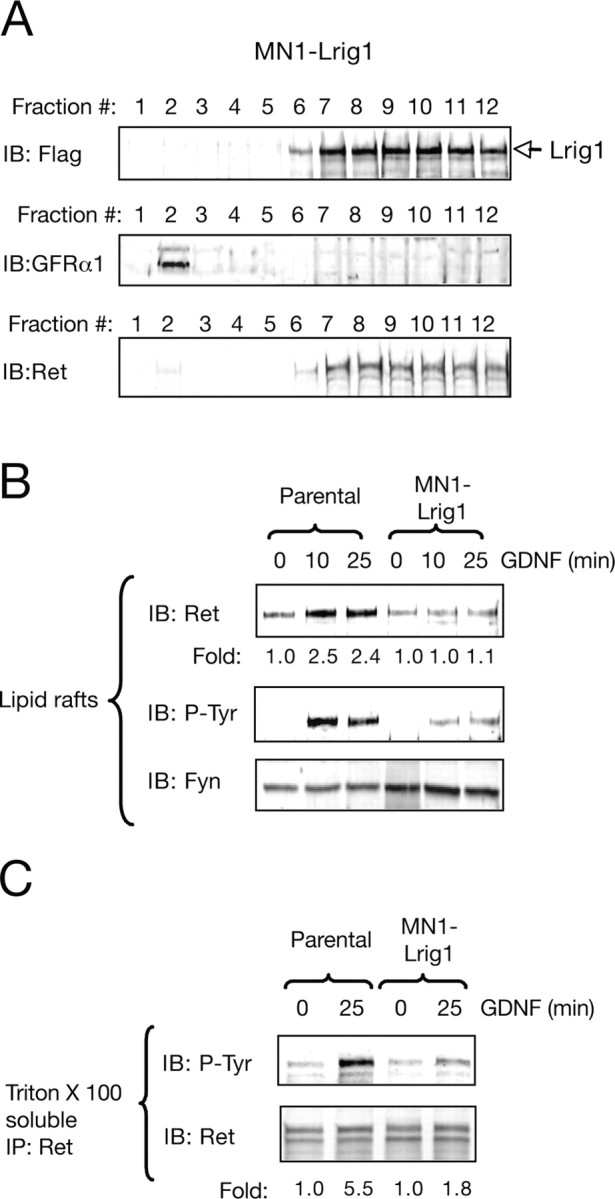
References
-
- Airaksinen MS, Titievsky A, Saarma M. GDNF family neurotrophic factor signalling: four masters, one servant? Mol Cell Neurosci. 1999;13:313–325. - PubMed
-
- Airaksinen MS, Saarma M. The GDNF family: signalling, biological functions and therapeutic value. Nat Rev Neurosci. 2002;3:383–394. - PubMed
-
- Arevalo JC, Waite J, Rajagopal R, Beyna M, Chen ZY, Lee FS, Chao MV. Cell survival through Trk neurotrophin receptors is differentially regulated by ubiquitination. Neuron. 2006;50:549–559. - PubMed
-
- Basson MA, Akbulut S, Watson-Johnson J, Simon R, Carroll TJ, Shakya R, Gross I, Martin GR, Lufkin T, McMahon AP, Wilson PD, Costantini FD, Mason IJ, Licht JD. Sprouty1 is a critical regulator of GDNF/RET-mediated kidney induction. Dev Cell. 2005;8:229–239. - PubMed
-
- Chen Y, Aulia S, Li L, Tang BL. AMIGO and friends: an emerging family of brain-enriched, neuronal growth modulating, type I transmembrane proteins with leucine-rich repeats (LRR) and cell adhesion molecule motifs. Brain Res Rev. 2006;51:265–274. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous