

L1 interaction with ankyrin regulates mediolateral topography in the retinocollicular projection
- PMID: 18171935
- PMCID: PMC6671135
- DOI: 10.1523/JNEUROSCI.3573-07.2008
L1 interaction with ankyrin regulates mediolateral topography in the retinocollicular projection
Abstract
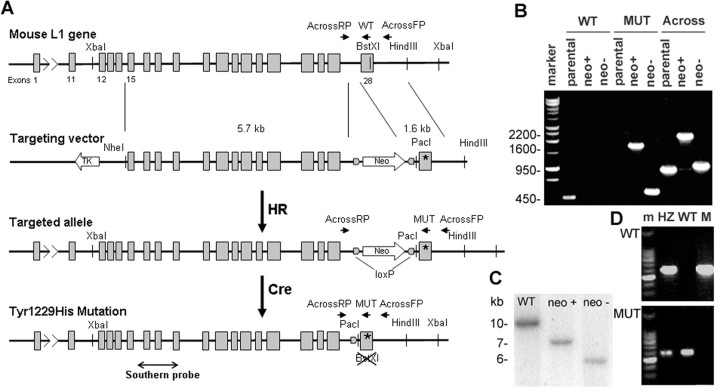
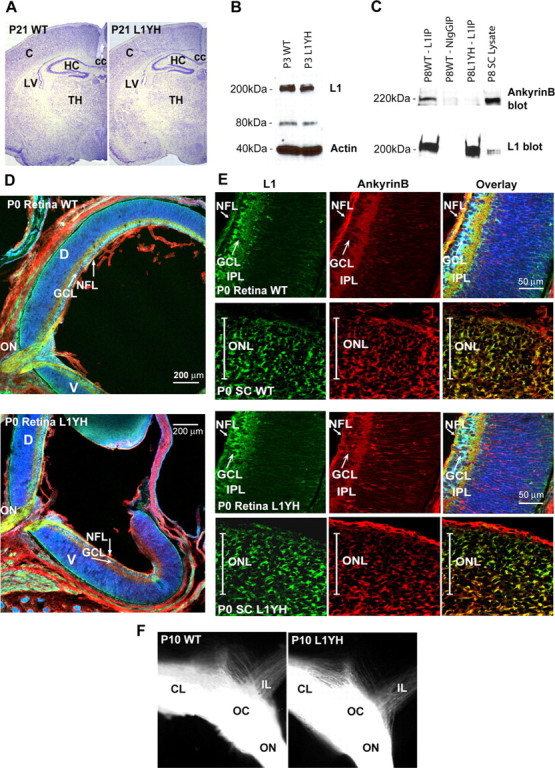
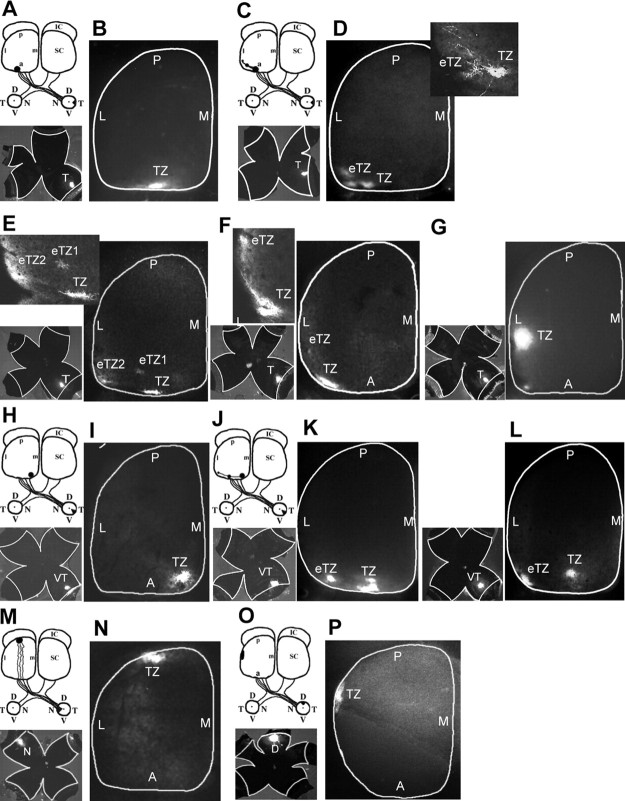
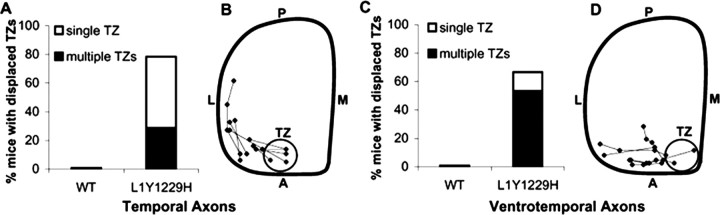
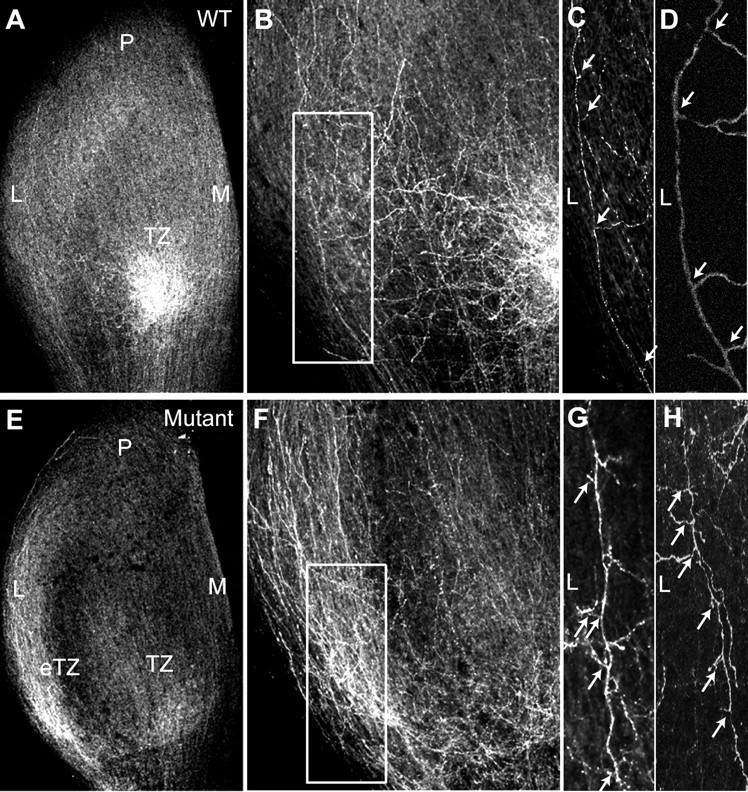
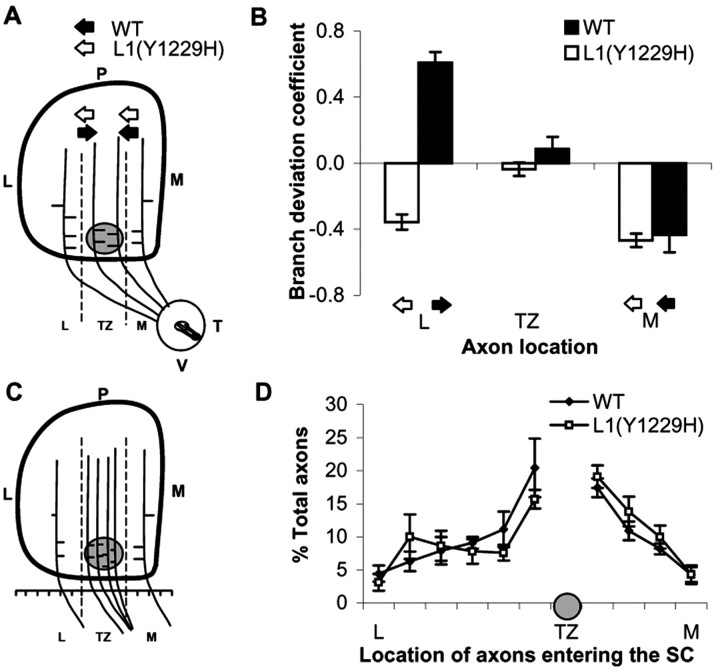
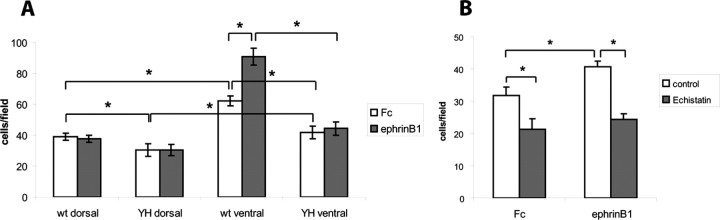
Dynamic modulation of adhesion provided by anchorage of axonal receptors with the cytoskeleton contributes to attractant or repellent responses that guide axons to topographic targets in the brain. The neural cell adhesion molecule L1 engages the spectrin-actin cytoskeleton through reversible linkage of its cytoplasmic domain to ankyrin. To investigate a role for L1 association with the cytoskeleton in topographic guidance of retinal axons to the superior colliculus, a novel mouse strain was generated by genetic knock-in that expresses an L1 point mutation (Tyr1229His) abolishing ankyrin binding. Axon tracing revealed a striking mistargeting of mutant ganglion cell axons from the ventral retina, which express high levels of ephrinB receptors, to abnormally lateral sites in the contralateral superior colliculus, where they formed multiple ectopic arborizations. These axons were compromised in extending interstitial branches in the medial direction, a normal response to the high medial to low lateral SC gradient of ephrinB1. Furthermore, ventral but not dorsal L1(Y1229H) retinal cells were impaired for ephrinB1-stimulated adhesion through beta1 integrins in culture. The retinocollicular phenotype of the L1(Tyr1229His) mutant provides the first evidence that L1 regulates topographic mapping of retinal axons through adhesion mediated by linkage to the actin cytoskeleton and functional interaction with the ephrinB/EphB targeting system.
Figures
References
-
- Ango F, di Cristo G, Higashiyama H, Bennett V, Wu P, Huang ZJ. Ankyrin-based subcellular gradient of neurofascin, an immunoglobulin family protein, directs GABAergic innervation at Purkinje axon initial segment. Cell. 2004;119:257–272. - PubMed
-
- Birgbauer E, Cowan CA, Sretavan DW, Henkemeyer M. Kinase independent function of EphB receptors in retinal axon pathfinding to the optic disc from dorsal but not ventral retina. Development. 2000;127:1231–1241. - PubMed
-
- Brakebusch C, Fassler R. beta 1 integrin function in vivo: adhesion, migration and more. Cancer Metastasis Rev. 2005;24:403–411. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases